Caveur à clé courte – Wikipedia
Le Cavalier à clé courte ( Macroscelides Prochoscideus ), parfois aussi comme Souris d’espionnage d’éléphants à oreille à courte Décrit, un type de mammifère du genre est Macroscélides et l’ordre des cavaliers du tronc (Macroscelidea). Il est répandu dans la partie sud-ouest de l’Afrique et vit dans les régions partiellement sèches et semi-désert du Karoo. Surtout, le museau de type tronc et le physique compact avec les membres minces sont caractéristiques. Dans le Karoo, le cavalier de lutte à courte durée habite terréstre comme un coureur rapide et utilise des zones d’incursion avec de nombreuses étagères naturelles. En tant qu’Omnivore, il détruit principalement les plantes et les insectes. Les animaux forment des liaisons monogames de couple, qui gardent principalement tout au long de la vie, les femelles donnent naissance à un ou deux garçons plusieurs fois par an. L’espèce a été décrite en 1800, il s’agit donc de la recherche la plus ancienne – représentative scénaristique de la proboscis. Surtout dans la seconde moitié du 20e siècle, le cavalier à clé courte était considéré comme le seul représentant du genre Macroscélides , Deux autres espèces sont décrites depuis 2012. L’inventaire n’est pas considéré comme menacé.


Le pull à tronc court est un petit représentant du cavalier de tronc et a un corps rond et une tête ronde. La longueur du fuselage de tête est de 10 à 11 cm, la longueur de la queue varie de 11 à 13 cm. Cela signifie que la queue est plus longue que le reste du corps. Le poids fluctue entre 31 et 47 g. La durée totale de cinq personnes de la Namibie examinée en juin 2007 se situait entre 22,4 et 23,6 cm, le poids était de 26 à 37,5 g. Avec les dimensions spécifiées, le cavalier à plantes à l’oreille courte représente l’un des plus petits types de famille, mais il est en moyenne plus grand que le cavalier de lutte à l’oreille courte Etendeka connexe ( Macroscelides micus ). [d’abord] Il n’y a pas de sexorphisme prononcé entre les sexes. [2] Le museau long et comme le tronc avec une grande vibration, qui atteint une longueur de 55 mm, est caractéristique. [3] L’oreille a une forme ronde et large et a des cheveux blancs blancs à l’intérieur. Comparé au cavalier de lutte à oreille court Etendka, le bord supérieur de l’oreille passe par-dessus la tête. Avec une longueur d’environ 21 à 29 mm, les oreilles ne sont pas particulièrement plus courtes qu’avec d’autres représentants de la proboscis. Les yeux atteignent une taille modérée, mais sont plus petites qu’avec les souris d’espionnage d’éléphant ( Éléphante ) et s’écarter de ces anneaux d’œil légers. La fourrure douce est colorée brun jaunâtre à gris sur le dessus, le dessous et les flancs sont plus brillants, par lequel la coloration ici peut passer de gris clair au blanchâtre. Les cheveux mesurent jusqu’à 17 mm de long et ont une base sombre. Des cheveux plus sombres et plus clairs se forment sur le dessus de la queue. À la fin, les cheveux deviennent plus longs, de sorte que la pointe a l’air légèrement touffue. Les glandes se produisent sur le dessous de la queue, qui ne sont pas toujours visibles par le cavalier à clé courte. S’ils semblent visibles à l’extérieur, ils atteignent une longueur de 8 à 12 mm, ce qui ne représente qu’environ 10% de la longueur totale de la queue. En comparaison, les glandes du cavalier de lutte à oreille court Etendeka prennent près d’un tiers de la longueur de la queue. La peau, qui est partiellement visible sous la couverture de fourrure, par exemple sur les oreilles, montre une pigmentation foncée. Dans ce document, le cavalier à clé à oreille est similaire au cavalier de rides namib Macroscelides flavicaudatus ), mais cela a une couleur de fourrure plus claire globale. Les minces membres postérieurs sont généralement beaucoup plus longs que les pattes avant. Les mains et les pieds ont chacun cinq rayons qui portent des griffes. Le pied arrière mesure entre 32 et 36 mm de longueur. [4] [5] [d’abord] [6]

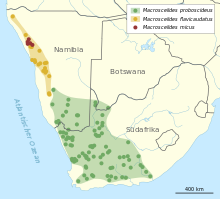
Le cavalier à clé courte est endémique en Afrique du Sud-Ouest. Il vit dans le sud et l’ouest de l’Afrique du Sud, dans le sud de la Namibie et dans la partie sud-ouest du Botswana. L’habitat comprend des régions sèches, principalement des déserts et des semi-déserts, principalement le karoo si appelé. Les niveaux de sable et de gravier dominent ici, mais sont souvent relativement densément couverts de buisson d’herbe et de végétation de la brousse. Les buissons peuvent mesurer jusqu’à 1 m de haut. Les températures annuelles moyennes dans le karoo fluctuent de 15 à 19 ° C, la précipitation annuelle est de 66 à 200 mm. La distribution de hauteur varie du niveau de la mer dans le succulent Caroo jusqu’à 1400 m dans le Nama-Karoo. [7] [5] L’ensemble de la zone de distribution est d’environ 500 000 km², peut-être plus. [8] La densité de population est considérée comme très faible. Après des enquêtes de 2005 à 2007 dans la réserve naturelle de Goegap en Afrique du Sud, elle a fluctué entre 0,35 et 1,59 individus par hectare. [2] Les deux autres représentants de Macroscélides Ayez une zone plus au nord, qui est largement située dans le Namib. Un couloir d’environ 50 km de large dans la réserve naturelle de Namibrand sépare la zone de distribution du cavalier à tronc d’oreille court de celui du cavalier de lutte à oreille à rude épreuve Namib. [7] [d’abord] [6]
Comportement territorial [ Modifier | Modifier le texte source ]]




Le pull à tronc court est un animal terrestre qui peut se déplacer très rapidement ( cursoriel ) et parfois jusqu’à 20 km / h devient rapide. Il vit à la fois la nuit et le crépuscule, les principales activités commencent vers 19h00 et se terminent tôt le matin vers 9h00. Les animaux sont rarement observés dans les heures de l’après-midi. [9] Les animaux individuels divertissent les salles d’action qu’ils utilisent sur une plus longue période de temps. Selon les enquêtes, la taille des zones de queue individuelles dans la réserve naturelle de Goegap est de 1,7 ha chez les hommes et 0,7 ha d’individus. Cependant, la taille augmente légèrement lors de l’élevage de jeunes animaux. La densité de la population représente un autre facteur d’influence. Dans les régions avec un faible nombre d’individus, les salles d’action des animaux mâles peuvent devenir significativement plus importantes et atteindre près de 3 hectares, mais cela a moins d’effet chez les femmes. Les maternels peuvent minimiser le risque d’être mis en proie. Malgré les dépenses parfois fluctuantes, les zones d’incursion des cavaliers courtes dans le karoo sont nettement plus petites qu’avec les parents du Namib, qui atteignent jusqu’à 100 ha. La fréquence des chevauchements des zones chez les individus du même sexe est très faible, comme cela est entre les deux sexes, ce qui suggère un certain degré de territorialité chez les hommes et les femmes. En général, les hommes et les femmes vivent dans des relations monogames en couple avec les pulls Trunks, qui peuvent durer jusqu’à la mort d’un partenaire. Cependant, les activités conjointes sont largement limitées au temps avant et pendant la ruée. Ensuite, le mâle reste près de la femme, le suit et superposé avec ses marques de parfums celles de son partenaire. Dans cette phase, les hommes défendent également le territoire de leur partenaire sur les autres couples vivant. En raison de ce comportement partiellement limité, le couple monogame se lie aux liaisons du tronc court est plutôt lâche, car les hommes visitent parfois deux femmes ou plus. Cette affaire se produit principalement lorsque des animaux féminins ont perdu leur partenaire. En règle générale, les mâles liés reviennent dans leur propre zone d’incursion quelque temps plus tard, principalement lorsqu’un mâle non lié suit également la femme sans partenaire. [2] [dix] Le cavalier à clé courte utilise diverses étagères distribuées dans les salles d’action individuelles. Les cachettes sont constituées de colonnes de roche ou de surplombs rocheuses, se trouvent sous les buissons et sous les buissons, les entrées sont souvent couvertes de végétation. Cependant, aucun nid spécial n’est créé dans les étagères. [4] [5] [11] [6]
Nourriture [ Modifier | Modifier le texte source ]]
Le cavalier à clé courte est un omnivore qui détruit à la fois des insectes et des matériaux à base de plantes. À partir d’un total de 46,5% d’insectes, la nourriture dans le karoo a en moyenne 48,4% sur les plantes, les 5% restants représentent les graines. Il est frappant que les femmes mangent généralement plus d’insectes que les mâles, ce qui peut être lié à la consommation d’énergie plus élevée et donc à la plus grande nécessité d’aliments riches en protéines pendant la grossesse et la phase d’allaitement des jeunes animaux. Cependant, il existe des différences régionales et saisonnières. Dans les zones occidentales du succulent Karoo, la proportion d’insectes et de plantes est plus ou moins la même au cours de l’année. Il existe des variations plus fortes dans les zones de distribution les plus orientales qui se caractérisent par deux saisons pluvieuses. La proportion annuelle d’insectes ici est de 63%, soit 36,7% sur les plantes. En été, jusqu’à 77,4% des insectes sont détruits, dans des cas individuels, même jusqu’à 88%, [douzième] [13] En revanche, seulement 42,5% en hiver. En cette période de l’année, contrairement aux zones occidentales, la consommation de plantes augmente car les insectes se produisent beaucoup moins fréquemment. Dans les examens comparatifs de plusieurs découvertes dans la zone de distribution, une variation individuelle élevée de la nutrition pourrait être déterminée globalement. Cependant, il faut souligner qu’une présence fréquente d’insectes n’augmente pas nécessairement la consommation de cela avec le cavalier de lutte à court terme. [14] [15] [16] [6]
la reproduction [ Modifier | Modifier le texte source ]]
La reproduction a lieu tout au long de l’année, l’oestrus des femmes utilise environ toutes les 10 semaines. La prévision des femmes peut souvent être observée en été (septembre à février), tandis que le nombre diminue considérablement au début de l’hiver de mars à juillet. [17] Selon les enquêtes de la réserve naturelle de Goegap, la durée moyenne de reproduction est donc un total d’environ huit mois par an pour chaque animal, tandis que les femmes reçoivent un total de deux à trois jeunes. Le style de vie monogame signifie que le mâle accompagne son partenaire principalement avant et pendant la ruée. Peu de temps avant que l’oestrus ne se produise, les animaux féminins frottent leur glande de parfum de la zone ano-génitale sur le terrain et mettent ainsi une sécrétion qui n’est couverte que peu de temps après par les propres marques de parfum du mâle, ce qui indique le partenariat existant. [dix] Le temps de soutien dure en moyenne 56 jours, pendant ce temps, le poids de la femelle augmente jusqu’à 20 g – un individu examiné pesait 64,3 g peu avant la naissance – ce qui correspond à environ la moitié du poids normal de la mère. [2] Après cela, la femelle donne généralement naissance à un ou deux jeunes animaux. En soins humains, la naissance de trois garçons pouvait très rarement être observée. [18] [3] La naissance a lieu dans un abri qui régule les taux de température entre le jour et la nuit. La construction des descendants est séparée de celle des parents masculins ou féminins, un nid spécial n’y est pas non plus installé. Les nouveau-nés sont déjà bien développés à la naissance, ils ont une fourrure douce et ont ouvert les yeux. Leur poids est d’environ 9 g. [3] Pendant l’élevage, il n’y a pas de soin direct des jeunes par le père animal, la mère animale ne rend visite aux garçons que irrégulièrement au ténien, en moyenne une fois par jour. Le temps de fusion est relativement court et prend environ deux à trois semaines. Les jeunes animaux mangent de la nourriture des insectes le cinquième jour. Ceci est initialement collecté et mâché par l’animal mère. À la fin de la période de fusion, les garçons commencent également à prendre des excursions plus importantes qui atteignent jusqu’à 240 m. Après environ six semaines, ils sont sexuellement matures et indépendants. [19] Étant donné que l’oestrus utilise quelques jours après la naissance de la progéniture, la femelle peut toujours transporter une nouvelle progéniture tout en élevant les jeunes. [3] [4] [5] L’espérance de vie dans les grands espaces est inconnue, mais il est supposé qu’il est de un à deux ans, similaire aux souris d’espion en éléphants liées. Jusqu’à huit ans et huit mois étaient en soins humains. [20] [21] [6]
Madorals et parasites [ Modifier | Modifier le texte source ]]
L’un des prédateurs les plus importants est la chouette d’orge, dans les restes voûtés du cavalier de lutte court. En cas de danger, un animal coule de Bush à Bush et, si nécessaire, se cache dans un abri. [4] [5] Les puces se produisent sous forme de parasites externes, les genres sont importants Échidnophaga et Xenopsylla . De plus, les tiques ont également été prouvées, notamment Rhicecephalus et Hémaphysalis . [22]
|
Systématique intérieure des cavaliers du tronc selon Heritage et al. 2020 [23]
|
Le cavalier à clé courte est une espèce du genre Macroscélides . Un total de trois types sont attribués à ce genre aujourd’hui, avec deux types uniquement érigés après 2010. Macroscélides Fait partie de l’ordre des cavaliers du tronc (Macroscelidea). Ce groupe de petits mammifères, uniquement en Afrique, est divisé en deux familles. [24] Pour les Rhynchocyonidae, seuls les chiens de tronc ( Rhynchocyon ) référé, ils sont monotypiques. Les chiens de tronc, qui sont également les plus grands représentants des pulls à coffre, vivent principalement dans des habitats. Les Macroscelididae forment la deuxième famille, il lui appartient à côté de Macroscélides Aussi les souris d’espion d’éléphant ( Éléphante ), les types du genre Galeshe , le rat de tronc ( Pétrodromus ) et la souris espion des éléphants nord-africains ( Pétaltator ) à. Tous les représentants des Macroscelidinae sont adaptés à des paysages ouverts significativement plus sèches jusqu’à des régions parfois en forme de désert. Les examens génétiques moléculaires ont montré une relation plus détaillée entre Macroscélides avec Galeshe , Pétrodromus et Pétaltator . Les deux familles se sont séparées de la ligne des ancêtres communs dans le bas Oligocène il y a environ 32,8 millions d’années, et une plus grande diversification des Macroscelidae a eu lieu de l’oligozène supérieur il y a environ 28,5 millions d’années. Macroscélides Le bas Miocène s’est formé il y a environ 19,1 millions d’années. [25] [26] [27]
|
Système intérieur de Macroscélides nach Dumbacher et al. 2014 [d’abord]
|


L’histoire taxonomique du genre Macroscélides est complexe. Dans la première moitié du XXe siècle, une division des cavaliers à courts métrages courts de deux manières avec jusqu’à 10 sous-espèces a été utilisée. Près de M. Proboscideus Également appliqué M. Melanotis En tant qu’espèces reconnues. Une révision du genre en 1968 M. Proboscideus Seule une manière valide contenait deux sous-espèces. Si limité lui-même M. p. proboscideus Sur les zones de carooine en Afrique du Sud, caractérisées par des tons de paysage plus sombres et plus affectés par les ombres. M. p. flavicaudatus En revanche, les régions plus légères et plus ensoleillées du Namib vivaient plus au nord. Les dessins en fourrure partiellement variés ont été considérés comme une adaptation aux conditions de l’habitat local. Jusqu’au début du troisième millénaire, le cavalier à clé courte était le seul moyen du genre Macroscélides . Cependant, l’occurrence et le nombre de sous-espèces ont été discutés. [28] [5] Ce n’est qu’au début des examens génétiques moléculaires du 21e siècle combinés à des recherches sur le terrain sur les preuves du site d’une division plus différenciée du genre. Les analyses génétiques ont pu séparer une population nord et sud, qui a également été confirmée par les examens de banlieue qui ont entraîné une séparation spatiale des deux groupes. En raison de l’isolement génétique également éprouvé des deux groupes, les chercheurs ont soulevé la sous-espèce en 2012 M. p. flavicaudatus Dans le statut d’art, avec lequel le cavalier à clé à oreille courte namib ( Macroscelides flavicaudatus ) En plus du cavalier à clé courte, la deuxième espèce reconnue du genre Macroscélides formé. Avec le cavalier de lutte à oreille court Etendka ( Macroscelides micus ) Deux ans plus tard, un troisième, aimable à la région, a été ajouté au cavalier à clé à l’oreille namib-short. En raison de la réévaluation de M. p. flavicaudatus En tant qu’espèce indépendante, aucune sous-espèce du cavalier à tronc court ne se distingue aujourd’hui. [7] [d’abord]
La première description scientifique du cavalier à clé courte a eu lieu en 1800 par George Shaw. Elle a eu lieu sous le type de nom Sorex Prochoscideus , avec lequel Shaw a mis le chemin des souris Rotzahnspitz. Shaw a nommé la couleur brune de la fourrure et le nez étonnamment long comme une plaque d’immatriculation spéciale. En tant que zone de type présumé, il a déclaré le Cap de Bonne-Espérance, [29] Ce qui a été spécifié en 1951 par Austin Roberts à Roodewal, Oudtshoorn, Cape Province (maintenant Westkap). Le cavalier à clé courte est donc le membre le plus décrit des cavaliers du tronc décrit dans l’histoire. [4] [5]
Malgré la zone relativement petite que le cavalier à clé courte habitée, aucune menace majeure n’est connue pour l’inventaire. Local, en particulier près de la rivière, le paysage à travers une agriculture à petite échelle ou industrielle, ainsi que l’expansion des centres de colonie urbaine. La formation ou la bague du désert associée peut également avoir un impact négatif sur les habitats du cavalier à tronc court, mais ces changements ne semblent actuellement que faibles. Pour cette raison, l’UICN considère l’inventaire comme «non en danger» ( le moins préoccupant ) un. [8]
Le cavalier de lutte à court terme est souvent présent dans les institutions zoologiques, en particulier en Europe en raison des coûts d’hébergement bas, de la forte attractivité et des séquences de reproduction souvent fréquentes. Depuis les années 1990, il y a eu en moyenne 24 à 25 propriétaires avec un total d’environ 100 animaux en Allemagne seulement. [30] [trente et un] L’une des races les plus importantes est dans le zoo de Wuppertal, [3] Un autre minis dans le zoo du zoo des minis dans l’erzgebirge. [30]
- John P. Dumbacher, Galen B. Rathbun, Timothy O. Osborne, Michael Griffin et Seth J. Eiseb: Une nouvelle espèce de Sengi rond (Genre Macroscelides) de Namibie. Journal of Mammalogy 95 (3), 2014, S. 443–454
- Stephen Heritage: Macroscelidae (je suis). Dans: Don E. Wilson et Russell A. Mittermeier (éd.): Manuel des mammifères du monde. Volume 8: Insectivores, paresseux et colugos. Lynx Edicions, Barcelone 2018, S. 206–234 (S. 229) ISBN 978-84-16728-08-4
- Ronald M. Nowak: Les mammifères du monde de Walker . Johns Hopkins University Press, 1999 ISBN 0-8018-5789-9
- Mike Perrin et Galen B. Rathbun: Macroscelides Prochoscide Sengi rond à oreilles rondes (époussette d’éléphants à oreilles rondes). Dans: Jonathan Kingdon, David Happold, Michael Hoffmann, Thomas Butynski, Meredith Happold et Jan Kalina (éd.): Mammals of Africa Volume I. Chapitres d’introduction et Afrotheria. Bloomsbury, Londres, 2013, S. 277–278
- Galen B. Rathbun: Macroscelides Proboscideus (Shaw, 1800) – Elephant à oreilles rondes. Dans: John D. Skinner et Christian T. Chimimba (éd.): Les mammifères de la sous-région d’Afrique australe. Cambridge University Press, 2005, S. 25-27
- Don E. Wilson et Deeann M. Reeder (éd.): Espèces de mammifères du monde . 3e édition. The Johns Hopkins University Press, Baltimore 2005, ISBN 0-8018-8221-4
- ↑ un b c d C’est John P. Dumbacher, Galen B. Rathbun, Timothy O. Osborne, Michael Griffin et Seth J. Eiseb: Une nouvelle espèce de Sengi rond (Genre Macroscelides) de Namibie. Journal of Mammalogy 95 (3), 2014, S. 443–454
- ↑ un b c d Melanie Schubert, Neville Pillay, David O. Ribble et Carsten Schradin: Le Sengi rond et l’évolution de la monogamie sociale: facteurs qui contraignent les mâles à vivre avec une seule femme. Ethologie 115, 2009, S. 972–985
- ↑ un b c d C’est Gea Olbricht et Alexander Sliwa: Éléphants souris Spitz – Les petits parents des éléphants? Journal of Cologne Zoo 53 (3), 2010, pp. 135–147
- ↑ un b c d C’est Galen B. Rathbun: Macroscelides Proboscideus (Shaw, 1800) – Elephant à oreilles rondes. Dans: John D. Skinner et Christian T. Chimimba (éd.): Les mammifères de la sous-région d’Afrique australe. Cambridge University Press, 2005, S. 25-27
- ↑ un b c d C’est F g Mike Perrin et Galen B. Rathbun: Macroscelides Prochoscide Sengi rond à oreilles rondes (époussette d’éléphants à oreilles rondes). Dans: Jonathan Kingdon, David Happold, Michael Hoffmann, Thomas Butynski, Meredith Happold et Jan Kalina (éd.): Mammals of Africa Volume I. Chapitres d’introduction et Afrotheria. Bloomsbury, Londres, 2013, S. 277–278
- ↑ un b c d C’est Stephen Heritage: Macroscelidae (je suis). Dans: Don E. Wilson et Russell A. Mittermeier (éd.): Manuel des mammifères du monde. Volume 8: Insectivores, paresseux et colugos. Lynx Edicionens, Barcelone 2018, S. 206-234 (S. 229-230) ISBN 978-84-16728-08-4
- ↑ un b c John P. Dumbacher, Galen B. Rathbun, Hanneline A. Smit et Seth J. Eiseb: Phylogénie et taxonomie des sengis à oreilles rondes ou des effets d’éléphants, le genre Macroscelides (Mammalia, Afrotheria, Macroscelidea). PLOS ONE 7 (3), 2012, S. E32410
- ↑ un b Galen B. Rathbun et Hanneline Smit-Robinson: Macroscelides Proboscideus. La liste rouge de l’UICN d’espèces menacées. Version 2015.2. ( [d’abord] )); Dernière consulté le 1er juillet 2015
- ↑ Lizanne Roxburgh et M. R. Perrin: Régulation et modèle d’activité de la température de la musaraigne d’éléphant à oreilles rondes (Macroscelides proboscide. Journal of Thermal Biology 19 (1), 1994, S. 13-20
- ↑ un b Melanie Schubert, Carsten Schradin, Heiko G. Rödel, Neville Pillay et David O. Ribble: Un compagnon mâle gardant dans un mammifère socialement monogame, le Sengi rond: sur les coûts et les compromis. Écologie comportementale et sociobiologie 64, 2009, S. 257–264
- ↑ Galen B. Rathbun: Pourquoi y a-t-il une diversité discordante dans Sengi (Mammalia: Afrotheria: Macroscelidea) Taxonomie et écologie? African Journal of Ecology 47, 2009, S. 1–13
- ↑ Graham I. H. Kerley: Consommation de graines de mammifères dans le Karoo, Afrique du Sud: preuve supplémentaire de divergence dans les processus biotiques du désert. Écologie 89, 1992, S. 471-475
- ↑ Graham I. H. Kerley: Statut trophique des petits mammifères dans le Karoo semi-aride, Afrique du Sud. Journal of Zoology 226, 1992, S. 563–572
- ↑ Graham I. H. Kerley: Régime des petits mammifères du Karoo, en Afrique du Sud. South African Journal of Wildlife Research 19, 1989, S. 67–72
- ↑ M. J. Lawes und M. R. Perrin: Comportement de nourriture sensible au risque de la musaraigne d’éléphant à oreilles rondes (Macroscelides proboscide). Écologie comportementale et sociobiologie 37, 1995, S. 31–37
- ↑ Graham I. H. Kerley: La courbe d’éléphant à oreilles rondes Macroscelides Proboscide (Macroscelidea) comme omnivore. Mammal Review 25, 1995, S. 39–44
- ↑ R. T. F. Bernard, G. I. H. Kerley, T. Doubell und A. Davison: Reproduction dans la musaraigne d’éléphant à oreilles rondes (Macroscelides Proboscide) dans le sud du Karoo, en Afrique du Sud. Journal of Zoology 240, 1996, S. 233–243
- ↑ Gea Olbricht, C. Kern et G. Vakhrusheva: Certains aspects de la biologie reproductive des trunkers à oreilles courtes (Macroscelides proboscide A. Smith, 1829) dans les jardins zoologiques avec une considération particulière des lancers de triplet. The Zoological Garden 75, 2005, pp. 304–316
- ↑ E. G. Sauer: Pour le comportement social des macroscelides de souris Elephant Spy Macroscelides Proboscide. Journal of Mammalkunde 38, 1973, pp. 65–97
- ↑ Richard Weigl: Longévité des mammifères en captivité; des collections vivantes du monde. Small Senckenberg Series 48, Stuttgart, 2005, ISBN 3-510-61379-1
- ↑ Gea olbricht: Longévité et fécondité dans Sengis (Macroscelidea). Afrotherian Conservation 5, 2007, S. 3–5
- ↑ L. J. Fourie, J. S. du Toit, D. J. Kok und I. G. Horak: Parasites d’arthropodes des effets d’éléphants, avec une référence particulière des tiques. Mammal Review 25, 1995, S. 31–37
- ↑ Sterith Eritage, Houseinin Rayaleh, le chef de la foule A.A Labu Kind. De nouveaux enregistrements d’une espèce perdue et d’une expansion de la gamme géographique pour Sengis dans la corne de l’Afrique. Peerj 8, 2020, S. E9652, Deux: 10.7717 / peerj.9652
- ↑ Brigitte Senut et Martin Pickford: Les mammifères micro-cursoriels de la fin de l’Éocène Tufas à Eocliff, Namibie. Communications of the Geological Survey of Namibia 23, 2021, S. 90–160
- ↑ Hanneline Adri Smit, Bettine Jansen van Vuuren, P. C. M. O’Brien, M. Ferguson-Smith, F. Yang et T. J. Robinson: Relations phylogénétiques des effets d’éléphants (Afrotheria, Macroscelidae). Journal of Zoology 284, 2011, S. 133–143
- ↑ Mike Perrin et Galen B. Rathbun: Ordonnez Macroscelidea – Sengis (Elephant-Erews). Dans: Jonathan Kingdon, David Happold, Michael Hoffmann, Thomas Butynski, Meredith Happold et Jan Kalina (éd.): Mammals of Africa Volume I. Chapitres d’introduction et Afrotheria. Bloomsbury, Londres, 2013, S. 258–260
- ↑ John P. Dumbacher, Elizabeth J. Carlen et Galen B. Rathbun: Petrosaltator Gen. Nov., Un nouveau genre remplace le Sengi nord-africain Elephantulus rozeti (Macroscelidea; Macroscelidae). Zootaxa 4136 (3), 2016, S. 567-579
- ↑ Don E. Wilson et Deeann M. Reeder (éd.): Espèces de mammifères du monde . 3e édition. The Johns Hopkins University Press, Baltimore 2005, ISBN 0-8018-8221-4, p. 472
- ↑ George Shaw: Zoologie générale ou histoire naturelle systématique. Volume I Partie II. Mammalie. Londres, 1800, S. 249–552 (S. 536) ( [2] )
- ↑ un b Liste des animaux du zoo ( [3] ), consulté pour la dernière fois le 16 octobre 2018
- ↑ Galen B. Rathbun et Laurie Bingaman Lacke: Une brève histoire graphique de Sengis en captivité. Afrotherian Conservation 5, 2007, S. 7–8
Recent Comments