Zapfen (Auge) – Wikipedia
Robinet ou Appuyer sur la cellule , anatomique Neurone coniferum [d’abord] (Latin conifère ‘Coat of Cones’), est appelé une sorte de récepteurs photo dans la rétine de la libe vertébrée avec un processus conique, le Robinet , anatomique RETINE CONE (Latin conus ,Cône’). Les cellules papales sont des neurones qui servent de cellules sensorielles spécialisées dans la vision photopique en lumière du jour et sont nécessaires à la perception des couleurs.
Chez l’homme, trois types de robinets sont différenciés, qui S -, M – et L -Casquette. Votre réponse de stimulus décrit la courbe d’absorption spectrale, qui forme la base de la métrique couleur.
Il y a des vertébrés avec un, deux, trois, quatre ou cinq types de robinets.
Le patch est un type de cellules sensibles à la lumière dans la rétine de l’œil. Ce sont des récepteurs photo des vertébrés qui permettent de voir avec les baguettes. Les patchs ne sont actifs qu’avec un éclairage suffisant car ils ne sont pas très sensibles à la lumière. Étant donné qu’au moins deux types de TAP de sensibilité spectrale différente se produisent chez la plupart des vertébrés, la perception des couleurs est généralement possible. L’homme a trois types de cônes différents, dont les maxima d’absorption sont d’environ 420 nm, 535 nm et 563 nm [2] [3] Ces longueurs d’onde correspondent aux couleurs bleues -violet ⬤ , Smaragdgrün ⬤ et vert jaune ⬤ . Le type de robinet moyen est évolutivement relativement nouveau et améliore la distinction entre les tons jaunes, rouges et verts.
L’activation prédominante de certains types de cônes se traduit par les couleurs correspondantes, mais les impressions de tous les types de TAP entraînent les impressions gris à blanc. Dans le cas d’un éclairance excessive – à la fois uniquement dans des endroits individuels comme un éclat ou dans tout le domaine de la vision – les cônes sont surfarés (saturés) et les aveugles se produisent.
Les baguettes sont saturées d’intensités lumineuses beaucoup plus faibles, en ce qui concerne la luminosité beaucoup plus sensible, c’est pourquoi elles ne contribuent pas à l’impression visuelle à la lumière du jour. Puisqu’il n’y a qu’un seul type de bâton chez l’homme, les baguettes ne peuvent pas entraîner une pression de couleur. Lorsque la lumière est diminuée, les cônes ne sont plus exposés suffisamment, ce qui signifie que les couleurs semblent disparaître, par exemple pendant le crépuscule. Avec la faible intensité d’exposition, seules les baguettes sont adéquatement irritées.
L’expression «cônes» a été utilisée différemment dans des contextes historiques et a décrit le staphylome sur l’œil.
Types de garniture d’homme [ Modifier | Modifier le texte source ]]
Il existe trois types différents de cônes chez l’homme.
-
- Scones
- S Récepteur de la longueur d’onde ou aussi k-cones ( k Longueurs d’onde d’Urze ). Ce récepteur couvre la zone bleue du spectre de couleurs visibles. Le maximum d’absorption est une longueur d’onde d’environ 420 nm (bleu-violet). [2] [3] Les deux informations se trouvent dans la zone de la lumière bleue, donc on l’appelle Blaurececepteur . Les Contes S de l’homme sont génétiquement étroitement liées aux cônes UV d’autres vertébrés. On parle donc de Cônes S2 -E contraste avec les S-Cones dans d’autres vertébrés.
Le blaurececepteur est rarement affecté par la couleur, mais s’il n’existe pas, l’amétropie survient. Par conséquent, la désignation obsolète remue T .
-
- M-CONNE
- M récepteur de la longueur d’ondium . Le maximum d’absorption du Récepteur vert est d’environ 534 nm (vert émeraude), [2] [3] Il couvre une zone entre la lumière bleue et orange. Les m-coules humains sont génétiquement étroitement liés aux L-Cones. On suppose qu’il n’a développé qu’il y a quelques millions d’années seulement par Genduplication des L-Cones. Les gènes de l’opsine respective des cônes L et M sont sur le chromosome X d’affilée.
S’il n’y a pas de récepteurs verts, l’amétropie survient. Par conséquent, la désignation obsolète remue C .
-
- L
- L Récepteur de longueur d’onde . Son maximum d’absorption est d’environ 563 nm (vert jaune). [2] [3] Néanmoins il sera aussi Rocher Décrit parce qu’il reprend la performance principale pour la perception de la lumière rouge. La broche L humaine est phylogénétiquement ancienne et correspond à tous les autres vertébrés. Le gène de son opsin est localisé sur le chromosome X.
S’il n’y a pas de récepteurs rouges, le sacrifice de la protanopie survient. Par conséquent, la désignation obsolète remue P .
Nombre de types de robinets d’animaux différents [ Modifier | Modifier le texte source ]]
-
- 0 types de taps
- Au moins deux espèces de primates nocturnes n’ont pas de cônes fonctionnels; Vous ne voyez pas de couleurs, juste un noir. [4]
-
- 1 type de robinet
- Certains mammifères tels que les phoques et la plupart des baleines sont des monochromates; Vous ne pouvez pas distinguer les couleurs.
-
- 2 types de robinets
- La plupart des mammifères (comme les chats et les chiens) n’ont que deux types de cônes (couleurs dichromatiques); Les M-Cones ne sont pas disponibles, ce qui est comparable à la cécité-vert rouge. [5] Cependant, les gouttelettes d’huile qui sont courantes dans les vertébrés actifs du jour ainsi que les doubles épingles principalement existantes se produisent toujours avec des animaux de toilette (monotrems) et des prières (Marsupialier), mais sont manquantes lors du placent.
-
- 3 types de robinets
- Les gens, l’ancien monde tandis que le personnel du Nouveau Monde ont généralement trois types de bandes; Vous voyez trichromatique. [5]
-
- 4 types de robinets
- Le plan du plan tétrachromatique des vertébrés à l’origine soupçonné contient quatre types de robinets différents [5] : Cônes UV, S-, M et L, dans lesquels les longueurs d’onde du maximum d’absorption respective (dans l’ordre mentionné) sont de 90 à 100 nm. De même, de nombreux arithts voient ainsi que des insectes et des araignées sauter ainsi que de nombreux oiseaux [6] Tétrachromatique. Environ douze pour cent de toutes les femmes européennes sont des tétrachromates avec des cônes L ou M anormaux supplémentaires, mais seuls quelques-uns peuvent l’utiliser pour une coloration plus précise. [6] [7]
-
- 12 types de robinets
- Le crabe de capture Neogondodactylus transfertii a huit types de recettes différents dans le visible et quatre dans la zone UV. [8]

La courbe d’absorption d’un type de robinet dépend de la construction de l’opsine de son pigment visuel, l’iodopsine. La transduction photochimique, c’est-à-dire la conversion des signaux de lumière en informations neuronales, fonctionne très également dans les baguettes et les cônes et les mêmes dans les types de robinets. La réaction d’un type de TAP à une certaine teneur spectrale de la lumière est donc déterminée par son type d’opsine.
Une somme pondérée des courbes d’absorption qui déterminent les récepteurs décrivent le Courbe de sensibilité spectrale Pour la vision quotidienne (le V (λ) -kurve ), son maximum est de 555 nm, ce qui correspond au vert moyen dans la lumière monochromatique de la couleur [9] (La valeur sert également à définir les candela et les unités dérivées telles que Lux). Le poids des trois courbes décrit les pondérés et moyennés pour l’observateur standard.
Le maximum pour les nuits des baguettes, en revanche, est de 498 nm (cyan). [2]
La commutation des cônes dans l’œil humain illustre le graphique suivant:


Dans le calque de photoecepteur ( Stratum neuroépithéliale ) La rétine humaine est de 6 millions de robinets et environ 120 millions de baguettes. [dix] [11]
La proportion de cônes sensibles au bleu chez toutes les personnes est presque constant de douze pour cent. Le rapport des cônes rouges et verts sur la rétine varie très fortement au sein d’une famille. La densité des cônes varie entre les espèces. Chez l’homme, la densité des cônes de la rétine dans le centre, la fovea centralis ou le «sehgrube», la zone de vision la plus forte, est la plus grande et diminue vers la périphérie. À l’inverse, la densité des baguettes augmente du centre à la périphérie. La différenciation des bâtons et des cônes a des raisons fonctionnelles: les cônes ne fonctionnent qu’à la luminosité et au crépuscule et rendent la vision des couleurs possible, tandis que dans des boulettes sombres ou avec une obscurité presque complète en raison de leur sensibilité beaucoup plus élevée à la lumière, seuls les baguettes fonctionnent. Les bâtons sont même capables de percevoir des photons individuels dans l’obscurité absolue, par laquelle cette perception peut être considérablement perturbée par les réactions spontanées à la chaleur, à la pression intraoculaire ou aux champs magnétiques très forts.
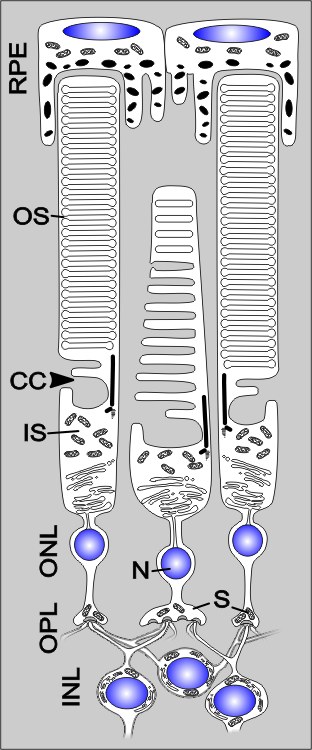
Sous la structure, les deux types de récepteurs photo – les cônes et les baguettes – sont organisés de manière similaire et consistent en un corps cellulaire, une synapse et une spécialisation cellulaire: le segment intérieur et extérieur.
Cependant, il y a des différences:
- Les cônes sont beaucoup plus larges que les baguettes.
- Dans les deux types de cellules, dans le segment externe (” Segment extérieur «, OS) La phototransduction au moyen de la protéine membranaire à sept trans opsine couplée à la rétine. Le pigment (opsine plus rétinien) est appelé iodopsine dans les robinets et est stocké dans de nombreux plis membranaires. Dans le cas des baguettes, il est analogue à la rhodopsine similaire d’iodopsine (“sehpurpur”) dans des “disques” si appelés.
- Les segments externes des cônes sont plus courts et doivent être contactés par l’épithélium pigmentaire rétinien (RPE) par des contreforts étendus en forme de chiffon, contrairement aux baguettes externes à baguettes.
Un segment extérieur est connecté au segment intérieur via un cil modifié dans un emplacement décentralisé, le zilium de connexion (“CILIUM de connexion”, CC). Neuf Microtubuli Dupletts dans une arrangement non onglet forment la structure intérieure de ce cil immobile.
Le segment intérieur métabolique actif se termine à cela (” Segment intérieur “, Est). À son tour, cela peut être divisé en ellipsoïde riche en mitochondries et au myoïde, qui contient le réticulum endoplasmique (ER). La biosynthèse des protéines a lieu ici.
La couche suivante est la couche externe de grains (” Couche nucléaire externe «, Onl), qui contiennent le noyau cellulaire avec le corps cellulaire. Ceci est suivi par la couche paciforme externe (” Couche plexiforme externe «, OPL) avec une région synaptique. Les synapses à l’extrémité proximale des récepteurs photo sont partiellement plates et partiellement réservées. Ces derniers sont des “synapses de ruban”, en termes de bande ou de structure en forme de plaque directement sur la zone active du présynapteur. De nombreuses vésicules synaptiques sont liées à la structure du ruban et un nombre beaucoup plus élevé de vésicules par période de temps peut être distribué par rapport aux synapses “normales”.
Dans l’obscurité, la membrane présynaptique du robinet (ou le bâton) est en continu le glutamate de neurotransmetteur. Si la lumière frappe sur les cônes, les canaux ioniques de sodium sont fermés dans la membrane des cellules du cône via une cascade de transduction du signal. Étant donné que le cône via son segment intérieur perd des ions de potassium en raison de la concentration élevée de potassium qui y prévaut et des canaux ioniques de potassium là-bas, il développe un potentiel récepteur négatif, i. c’est-à-dire qu’il hyperpolarise et libère moins ou plus de glutamate.
Le glutamate de neurotransmetteur peut avoir un effet passionnant ou inhibant sur les cellules bipolaires en aval, car il existe deux types différents de cellules bipolaires, ce que l’on appelle on-bipolaire et off-bipolaire. Si un cônes sur un bipolaire, la distribution réduite du glutamate provoque une dépolarisation du bipolaire lors de l’exposition. Le glutamat a un effet inhibiteur sur le bipolaire, donc l’inhibition ne s’applique pas en ce qui concerne l’exposition. Cet effet est basé sur le fait que les récepteurs du glutamotma appelé mGlur6 sont stockés dans la membrane post-synaptique du métabotrope bipolaire. Dans l’obscurité avec du glutamate, les récepteurs MGLUR6 activent une cascade de signal dans le bipolaire qui ferme les canaux cations, i. c’est-à-dire que la cellule devient inutile. Si le glutamate est manquant, les récepteurs MGLUR6 restent vacants, ouvrent les canaux de cation du bipolaire, ils dépolarisaient et transmettent l’excitation. Ce mécanisme converserait l’hyperpolarisation du côté de la bipolaire lorsque l’exposition dans les cônes dans les cônes, c’est-à-dire sur bipolaire est excitée lors de l’exposition et inhibée par l’assombrissement.
Le deuxième type, les Off-Bipolaires, réagissent lors de l’exposition et réduisait ainsi la libération glutamale des cônes avec hyperpolarisation. Ils ont des récepteurs de glutamate ionotropes qui ferment les canaux cations. Cela signifie que les offices sont inhibées par la lumière et excitées par l’assombrissement.
La séparation dans le classage ON et OFF-Bipolaire est conservée tout au long de la piste visuelle ultérieure du cerveau.
Le glutamat est en fait considéré comme un neurotransmetteur excitant typique. Ce système montre qu’en fin de compte, les récepteurs post-synaptiques du glutamate décident de l’excitation ou de l’inhibition.
Le système visuel des primates détecte différentes couleurs sur l’activation (ou l’hyperpolarisation) des cônes dans la rétine. Le duplexretina humain contient trois types de cônes, la sensibilité spectrale différente (mais chevauchant) est générée par trois types d’opsine différents (iodopsine). [douzième] Avec un système nerveux construit en conséquence, les pigments photo permettent un trichromatique (c’est-à-dire sur la combinaison de trois couleurs primaires). Le résultat est la capacité étonnante des humains à pouvoir distinguer environ deux millions de nuances de couleur.
La perception des couleurs des mammifères et donc aussi des humains, de grands singes et des primates diffère considérablement en ce qui concerne le nombre de pigments visuels et donc les nuances de couleur perceptible.
Seuls les primates de l’Asie et de l’Afrique (personnel du vieux monde), qui incluent les gens, ainsi que en partie les relais sud-américains du Nouveau Monde, ont atteint la capacité des couleurs améliorées au cours de leur évolution. La capacité de nombreux primates à distinguer le rouge du vert apporte parfois des avantages vitaux. Par exemple, les fruits matures colorés peuvent être reconnus plus rapidement entre les feuilles vertes ou les jeunes feuilles plus digestibles peuvent être distinguées de l’ancien digestible. Les feuilles riches en nutriments ont souvent une légère couleur rouge ainsi que de nombreux fruits matures qui peuvent même contenir des toxines au stade encore vert. C’est pourquoi cette capacité a prévalu dans l’évolution. La duplication d’un gène a été démontrée dans l’ancien monde où se trouve un peu plus que le long pigment visuel. En plus des informations sur le pigment bleu, le matériel génétique contenait également une vue rouge et sensible au vert. Les trois types de robinets avec leurs différents maxima d’absorption ont été créés dans de nombreux primates. [13]
La recherche a montré que le développement de la vision des couleurs est étroitement lié à une réduction de l’odeur, donc une partie de cette capacité a été perdue en faveur de la vision colorée chez ces primates. Les primates avec une meilleure perception des odeurs, en revanche, ont une capacité moins bien prononcée à distinguer la couleur. [14]
- Webvision : Aperçu de la structure et de la fonction des récepteurs photo et de la rétine (anglais)
- Couleurs Sur spectrum.de
- ↑ Terminologie histologique (Th, nomenclature actuelle), voir l’entrée H3.11.08.3.01046 S. 110 .
- ↑ un b c d C’est F J. K. Bowmaker, H. J. A. Dartnall: Pigments visuels de tiges et de cônes dans une rétine humaine. Dans: Le Journal of Physiology. Vol. 298, janvier 1980, ISSN 0022-3751 , S. 501–511, PMID 7359434 , (Fichier PDF; 1,5 Mo).
- ↑ un b c d Ralf Brandes, Florian Lang, Robert F. Schmidt: Physiologie de l’homme: avec physiopathologie . 32. Édition. Berlin; Heidelberg 2019, ISBN 978-3-662-56468-4, S. 759 , est ce que je: 10 1007 / 978-3-662-56468-4-4 .
- ↑ Gerald H. Jacobs, Maureen Neitz, Jay Neitz: Mutations dans les gènes pigmentaires de la cône et l’absence de vision des couleurs dans deux espèces de primate nocturne. ( Mémento à partir du 31 mai 2013 dans Archives Internet ) Dans: Actes de la Royal Society of London B: Sciences biologiques. Bande 263, Nr. 1371, 1996, S. 705-710.
- ↑ un b c La vision des couleurs des primates Sur primata.de, consulté le 13 mai 2014.
- ↑ un b Georg Rüschemeyer: Le quatrième cône. Dans: faz.net. à partir du 22 septembre 2012, consulté le 15 mars 2019
- ↑ Projet de tétrachromacy – La science. Université de Newcastle, Récupéré le 15 mai 2019 (Anglais).
- ↑ Le monde coloré des crabes. Dans: Spectre de la science. Janvier 2000, S. 13 , Récupéré le 27 novembre 2015 .
- ↑ Selon le cercle des couleurs d’Aemilius Müller
- ↑ Evelin Kirkilionis: Maladie de Minamata à la prédisposition (= Lexique de la biologie. Vol. 6). Herder, Freiburg (Breisgau) 1986, ISBN 3-451-19646-8, p. 158: Rétine.
- ↑ Neil A. Campbell: Biologie. Spectre, Akademischer Verlag, Heidelberg etc. 1997, ISBN 3-8274-0032-5, p. 1123 (indique 125 millions en tant que numéro de tige).
- ↑ Barry W. Connors, Michael A. Paradiso, Andreas K. Engel, Andreas Held, Monika Niehaus: Neuroscience un manuel de base pour la biologie, la médecine et la psychologie . 4e édition. Berlin, Allemagne 2018, ISBN 978-3-662-57262-7.
- ↑ Comment les ancêtres de l’homme ont appris la vision des couleurs Sur Farbimpulse.de, consulté le 13 mai 2014.
- ↑ Kathrin Gerewitz: Vision des couleurs au détriment de l’odeur Dans: Berliner Zeitung. à partir du 21 janvier 2004, consulté le 13 mai 2014.
Recent Comments