Canis Dirus -Wikipedia、無料百科事典
犬の恐怖 – 彼の競争相手の隣にあるセイバーの猫の隣に、最も有名な先史時代の北米の捕食者の一人である犬の家族からの哺乳類の絶滅種 スミロドン致命的 。彼は更新世後期(125〜1万年前)のアメリカ大陸に住んでいました。この種は、最初の標本の発見から4年後の1858年に命名されました。 2つの亜種があります。 C. Dirus Guildayi 私 C.と言うでしょう 。種はおそらくから来ています Dog Armbrusteri 。標本の最大のコレクション C.ドレッド 彼はカリフォルニア州ロサンゼルスのランチョラブレア出身です。
残っています C.ドレッド それらは、北米の乾燥したサバンナだけでなく、平原、草が茂った地域、北アメリカの山岳地帯など、幅広い生息地を表す地域で発見されました。彼らは最大2255 m A.S.Lをカバーしています。北の北に到達している地域は、さらに北に位置する地域に関する未確認の報告があります。範囲の北部の境界は、温度、獲物の利用可能性、生息地の利用可能性に関連しており、ローレン派の氷床とコルディーリー氷床の近さに起因しています。
C.ドレッド 多かれ少なかれ最大の現代的な灰色のオオカミの大きさに達しました、 カニス・ループス・パンガシレウス とカナダのオオカミ。 CD。ギルダイ 平均60 kgの重量、a CD。ひどい 平均して68 kg。頭蓋骨と歯は灰色のオオカミに似ていましたが、歯 C.ドレッド 彼らはより大きく、切断に適していましたが、動物はまた、現代のオオカミよりも牙の噛みつき力を持っていました。特徴は、後期 – オーティカンの大きな草食動物の狩猟への適応です。北米では、彼は狩りをしました:馬、怠zy、マストドントン、バイソン、絶滅、アメリカのタイプのラクダ カメロプ 。オオカミの他の大きなハイパーミアンを食べるように、彼はグループで狩りをしました。彼は、彼の獲物のジャンルとのクリーリーの絶滅の間に死にました。絶滅の原因として、メガロを食べる人への依存は、他の種の気候変動と競争とともに与えられましたが、それは議論の余地があります。最後の遺跡は9440年前から来ます。

1850年代から、米国では偉大なオオカミの化石の遺跡が見つかりました。それらが単一の種に属していることはすぐには説明されませんでした。後で割り当てられる最初の標本 犬の恐怖 、1854年の夏にインディアナ州エバンズビル近くのオハイオ川の入植地で発見されました。頬の歯のある化石顎は、フランシス・A・リンクのエバンズビルのコレクターからジョセフ・グランビル・ノーウッドという名前の地質学者の手に渡りました。古生物学者のジョセフ・ライディは、標本がオオカミの絶滅した種を表していると判断し、その名前で発見を記録しました Dog Primaevus [2] 。ライディへのノーウッドの手紙は、フィラデルフィアの自然科学アカデミーでホロタイプとともに保存されていました。 1857年、ネブラスカ州のニオブララ川渓谷の探検中に、レイダは絶滅した種のサークルを見つけました カニス 彼は次の年に名前で説明しました C.ドレッド [初め] 。名前があります C. Primaevus 後で変更された(Leidy、1854) 犬インディアンセンシス (Leidy、1869)、彼がその名前に気付いたとき C. Primaevus 以前はイギリスの自然主義者のブライアン・ホジソンによって使用されていました。 [3] 。
1876年、動物学者のジョエル・アサフ・アレンが遺体を発見した Dog Mississippiensis (1876年アレン)とそれらを関連付けました C.ドレッド (ライディ、1858)i 犬インディアンセンシス (Leidy、1869)。これらの3つの発見については、アレンがより多くの資料が見つかる前に、各標本を間に合わせの名前の下に残すのに最適だと考えていたので、彼らの親族を明らかにすることはほとんどありませんでした。 [4] 。 1908年、古生物学者のジョン・キャンベル・メリアムは、ランチョ・ラ・ブレアの大きなオオカミの洗練された骨の断片を数多く見つけ始めました。 1912年まで、彼はこれを正式に認識するのに十分なスケルトンを見つけ、以前は標本を種として見つけました C.ドレッド (Leidy、1858)。これまでに使用された中で最も古い名前を考慮して、命名法の原則のため [十] メリアムは1858年からライディの標本の名前を選びました。 C.ドレッド [11] 。 1915年、古生物学者のエドワード・トロクセルはメリアムに同意し、認識しました C. Indianensis 同義語の場合 C.ドレッド [12番目] 。 1918年、これらの化石を調べた後、メリアムはこれらの名前の組み合わせを別のタイプで提案しました エノシオン (から エノス 、ひどい、そして Cyon 、オオカミ)作成する エノシオンの恐怖 [6] 。しかし、誰もがこの絶滅したオオカミを別のタイプに配置することに同意したわけではありません。 [13] 。 アイリシ犬 (Sellards、1916)i エノシオンの恐怖 (Merriam、1918)は同義語として認識されました C.ドレッド 古生物学者アーネスト・ルンデリウス1972 [14] 。上記の税はすべて同義語として認識されていました C.ドレッド また、1979年に古生物学者のロナルド・M・ノバックの仕事で [15] 。
1984年の調査で、BjörnKurtenは人口内の地理的変動性を発見しました C.ドレッド 。彼は2つの亜種の区別を提案しました。 犬のディルス・ギルダイ (カリフォルニアとメキシコの標本のための古生物学者ジョン・E・ギルデイを記念してそう命名されました)、より短い手足と長い歯があり、 悲惨な犬 北米の水省の東側からの標本の場合、足が長く、歯が短い [8] [16] [17] [18] 。
進化 [ 編集 | コードを編集します ]
系統学 犬の恐怖 形態に基づいています [19] [20]
1974年、ロバートA.マーティンはグレートオオカミを渡すことを提案しました C. Armbrusteri 灰色のオオカミに C.ループス [21] 。 Nowak、Kurten、Annaisa Bertaはそれを決めました C.ドレッド 彼はこの灰色のオオカミから来ませんでした C.ループス [15] [22] [23] 。 1987年に、哺乳類の集団が英語と呼ばれるより大きな形を生み出すことができる新しい仮説が提起されました ハイパーモルフ 豊富な食べ物の間。食べ物が再び始まると、 ハイパーモルフ 適応したり、小さな形に移動したり、死にかけたりします。この仮説は、多くの後期哺乳類の大きな体の大きさが今日の親relativeと比較できることを説明することができます。絶滅と専門分野の両方が、極端な気候の時代に現れる可能性があります [24] [25] 。グロリア・D・グーレットはマーティンに同意しましたが、この仮説が急速な外観を説明できると提案しています C.ドレッド 北米では、彼らの頭蓋骨の類似性を評価する C.ループス Hypermorfを出産しました C.ドレッド 豊富な食物、環境の安定性、および大規模な競合他社の存在のため [26] 。
3人の古生物学者Xiaoming Wang、Richard H. Tedford、およびRonald M. Nowakがそれを提案しました C.ドレッド から進化しました Dog Armbrusteri [27] [20] 。ノバックは、メリーランド州のカンバーランド洞窟で見つかった標本はそうだと言った C. Armbrusteri 差別化 C.ドレッド [28] [29] 。中国からの初期のオオカミ、 カニス・アムリエンス 、両方の祖先であったかもしれません C. Armbrusteri そして灰色のオオカミ C.ループス [30] 。突然の外観 C. Armbrusteri 北米では、更新世の初期では、移民がアジアから来たことを示唆しています [最初に30] 、後半の更新世のように C.ループス [最初に30] [32] 。 2010年、Francisco Prevostiはそれを提案しました C.ドレッド 姉妹の分類群でした C.ループス [33] 。
犬の恐怖 彼は更新世後期から北米および中央アメリカにある中世前期(125000年から10000年前)に住んでいた [8] 。東部のほとんどが発見されています CD。ひどい 日付125,000〜75,000数年前、しかし西部の化石 CD。ギルダイ それらは小さいだけでなく、より現代的です。したがって、それはそれを提案されました CD。ギルダイ 彼はから来ました CD。ひどい [8] [18] 。ただし、議論の余地のある標本があります C.ドレッド 日付250,000年。化石 C.ドレッド シェリダン(ネブラスカ)の4つの場所で発見された干し草スプリングスエリアは名前が付けられました エノシオンの恐怖nebrascensis (フリック1930、言葉では言い表せない)が、その説明は公開されていません。 Nowakは後にこの資料を参照しました C. Armbrusteri [34] 。 2009年、テッドフォードは標本の説明を正式に公開し、その特徴的な形態学的特徴を提示したが、 C. Armbrusteri 、 私 C.ドレッド 、そのみを参照してください C.ドレッド [30] 。ネブラスカからのこれらの化石は、初期の化石記録を表すことができます C.ドレッド [7] [30] 。
サウスダコタ州のブラックヒルズの馬の部屋で発見された化石 C.ドレッド 。この場合、この化石はこの種の最も初期の標本の1つです [17] [35] 。としてカタログ化されています Cf.犬C.ドレッド [36] (どこ cf. ラテン語を意味します:議論、不確か)。馬の部屋で見つかった馬の化石は、252、000年でウラントラック法の年代測定を可能にしました。標本と考えられていました Cf.犬ひどい 同時に来る [17] [36] 。 C. Armbrusteri 私 C.ドレッド いくつかの一般的な機能(Synapomorph)を共有し、それを示唆しています C.ドレッド 約2、500年前に大陸の内部のオープンエリアで作成されました。 C. Armbrusteri [20] 。時間 C.ドレッド 25、000年前カリフォルニアとネブラスカで、そしてその後、今日の米国、カナダ、メキシコ、グアテマラ、ベネズエラ、エクアドル、ボリビア、ペルーの他の地域で来るでしょう [30] 、しかし、初期の化石のアイデンティティは確認されませんでした [37] 。
南アメリカの標本 C.ドレッド 更新世後期から西海岸に沿って発見されましたが、1つはアルゼンチンで発見されました。 カニスゲジ 私 カニス川 [38] 。ある研究によると C.ドレッド 最も進化した種でした カニス 新しい世界で、そしてと比較されます C.川 それは大きく、またより効率的な狩猟を可能にする低臼歯もありました [39] 。このため、一部の研究者はその論文を提供しました C.ドレッド 彼は南アメリカで作成されたかもしれません [40] [17] [23] 。 2009年にはそれが提案されました C. Armbrusteri 彼は北アメリカと南アメリカの共同祖先でした [38] 。翌年、この研究は、その結論につながる証拠を提供しました C.ドレッド 私 C.川 彼らは同じ種を構成したので、 C.ドレッド 北米から南に移住しました [33] 。
Perriと同僚(2021)は、個人の遺跡から得られたゲノムを選択しました C.ドレッド 50、000年から13、000年以上前の生活。系統発生の位置に関する著者の研究から C.ドレッド 灰色のオオカミとの解剖学的類似性にもかかわらず C.ドレッド 彼は彼と密接に関係しておらず、570万年前に、灰色のオオカミの祖先であるプラリーコヨーテを覆う進化ラインから分離された進化ラインに属していました。 ルパスター 、エチオピアのケーカー、ルディ・シオナ、リカオン・プストレゴ、サカリ。これを念頭に置いて、ペリと同僚は排除を推奨しました C.ドレッド 種類の カニス 別のタイプに転送します エノシオン [41] 。
系統樹は、ペリと同僚に基づいた生きている「オオカミ」犬の関係を描いています(数百万年の時間) [41] 。
DNA分析 [ 編集 | コードを編集します ]
1992年に、スケルトンの残留物からmtDNAを抽出しようとする試みがなされました CD。ギルダイ 他の種のオオカミとの関係を指定するための比較目的のため。遺体がラブレアの牧場から引き出され、タールを取り外すことは不可能だったため、試みは失敗しました [42] 。 2014年、コロンビアのマンモスのDNAはDNAを抽出しようと試みられず、研究者はアスファルトからの有機化合物がラブレアのすべての古い標本の骨に浸透し、DNA抽出を妨げるという結論に達しました。 [43] 。
放射性炭素年代測定 [ 編集 | コードを編集します ]
この種の遺跡のほとんどの場所の年齢は、生体序によってのみ決定されましたが、これはアスファルト堆積物の信頼できる指標ではありません [44] [45] 。一部の場所は、放射性炭素法を使用して日付が付けられていました。検体 C.ドレッド La Brea Pitsから、次のように:13000年から14000年前の82個の標本、14000年から16000年前の40個の標本、14000年から18000年あたり37個の標本、17000年から18000年に37個の標本、21000〜30000年の26個の標本、25000〜28000年あたり40個の標本、3000〜37000年の6個の標本 [46] 。ミズーリ州のパウダーミルクリーク洞窟の標本は、1999年に13170年に推定されました [17] 。

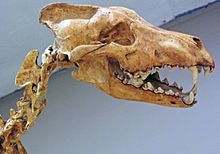
犬の恐怖 彼はオオカミのタイプの最も有名な科学代表でした [11] [27] 。体のシルエットとプロポーションは、2つの現代的な北米のオオカミに似ていました。 カニス・ループス・パンガシレウス [47] [11] 私 西オオカミ [47] 。最大の現代的な灰色のオオカミは、97 cm以下の高さに達し、体の長さは180 cm以下に達します [48] 。いくつかの標本 C.ドレッド ラブレア牧場では、彼らは与えられた次元に道を譲りましたが、他の人はそれらを超えました [11] 。
犬の恐怖 彼は、同じサイズのモダンな灰色のオオカミと比較して、足が小さく、大きな頭を持っていました。頭蓋骨の長さは、ユーコンのオオカミと比較して、より広い口蓋、額、ヨークで最大310 mm、さらにそれ以上に達する可能性があります。指定された寸法は、頭蓋骨の大量性を証明します。彼女のfi骨は高く、イニオンは後方向に注意を引くタブを着用しました。鼻骨の後端は頭蓋骨の後ろに到達しました。ただし、骨が多くの方向に分離して進むことができるため、Rancho La Breaに保存された骨接続を備えたスケルトンを見つけることは困難です。脊椎の一部が敷設されており、灰色のオオカミのホモロジストに似ているように見えます。椎骨の数は同じままです [11] 。
地理的な違い C.ドレッド スケルトンの残留検査により、頭蓋骨と歯のいくつかの特徴の違いと、カリフォルニアとメキシコの標本間の手足の割合が明らかになったとき、それは発見されませんでした(カリフォルニアとメキシコの標本の割合が明らかになりました。 CD。ギルダイ )そして、給水線の東側からの動物( CD。ひどい )。手足のサイズの比較により、後部の足が示されました CD。ギルダイ 脛骨と中足骨の骨がはるかに短くなったため、ジュコスキウルフよりも8%短かった。遠位骨が短いため、前肢も短かった [49] [50] 。比較的軽くて小さくて足と巨大な頭があります CD。ギルダイ 彼は灰色のオオカミやプレレリーコヨーのように走りに適していませんでした [11] [50] CD。ひどい 彼ははるかに長い後肢を持っていました CD。ギルダイ 。前肢はUよりも14%長かった CD。ギルダイ 上腕骨の10%、15%長いradial骨、15%長い金属。後肢はUより10%長かった CD。ギルダイ 大腿骨と脛骨が10%長く、中足骨が15%長くなっています。 CD。ひどい 手足の長さは、ジュコスキーウルフに似ています [49] 。最大の大腿骨 CD。ひどい ミズーリ州のキャロル洞窟で見つかり、278 mmを測定しました [18] 。

| 骨 | CD。ギルダイ [50] | ゆくウルフ [50] | CD。ひどい [49] |
|---|---|---|---|
| ブラジー | 218 mm | 237 mm | 240 mm |
| 半径 | 209 mm | 232 mm | 240 mm |
| メタカルパルの骨 | 88 mm | 101 mm | 101 mm |
| 大腿骨 | 242 mm | 251 mm | 266 mm |
| 脛骨骨 | 232 mm | 258 mm | 255 mm |
| 中足骨 | 93 mm | 109 mm | 107 mm |
質量 CD。ギルダイ 一方、60 kgと推定されます CD。ひどい 平均して68 kgで、一部の個人は大きかった [18] しかし、スケルトンの制限のため、それらは110 kgを超えませんでした [51] 。ジュコスキーウルフの平均体重は、男性の場合は43 kg、女性では37 kgであることを比較してください。ジュコスキウルフの個人の体重は21 kgから55 kgになります [52] 、個人の1人が79.4 kgに達しました [48] 。したがって、平均に続きます C.ドレッド サイズが最大のモダンな灰色のオオカミに似ていました [18] 。
完全な男性の遺物 C.ドレッド 彼の菌は彼をすべての犬と大幅に区別するので、時々他のオオカミと区別するのは簡単です [17] [49] 。

適応とは、特定の生息地の生物を生き残る能力が増加する進化プロセスを意味します [54] 。生息地、気候、獲物の種類、競争の種類などの生態学的要因は、灰色のオオカミの頭蓋骨と歯の可塑性に大きな影響を与えることが判明し、頭蓋骨と歯の適応の発達につながります [55] [56] [57] 。同様に C.ドレッド 彼は絶対的な肉食動物であり、頭蓋骨と歯科が大きくて抵抗する獲物での狩猟に適応していました [58] [59] [60] 。彼の頭蓋骨と口の形は時間とともに変化し、体の大きさの変化は気候変動と相関していました [六十一] 。
古生態学 [ 編集 | コードを編集します ]
最後の氷河期、125千人から続く。 [62] 最大14.5,000数年前 [63] 、更新世の終わりに、それは最新の口論です [62] 。氷の覆いが33,000人増加し始めた最後の氷河最大のピークに達しました。数年前、最大サイズの26.5,000に達しました。数年前。北半球では約19000人の脱尿が始まりました。数年前、南極では約14.5千人でした数年前、溶けた氷河からの水の証拠と一致しているのは、海面14.5千の急速な増加の主な原因としてです。数年前 [63] 。ウィスコンシンの氷河は北米にブロックされました。大陸の多くの場所からの化石の証拠は、最後の氷河期の終わりに、メガファウナと呼ばれる多くの大型動物の絶滅を示しています [六十四] 。
60,000から南カリフォルニア沿岸数年前、最後の氷河最大の終わりまで、それは今日よりも湿度に関しても涼しく、それ以上でした。最後の氷河最大時には、年間平均気温は11°Cから5°Cに低下し、年間降雨量は100から45 cmに減少しました。 [65] 。しかし、この地域はウィスコンシンの氷河作用によって過小評価されていましたが、動物と冷たい感受性の植物の避難所と考えられています [66] [六十七] [68] 。 24000数年前、茂みとオークの森とチャパラルの領域が減少しましたが、松の参加は増加しました。ジュニパーを備えた現代の太平洋山の森を連想させる長い森林が作成されました。 14000人の後BPの年、針葉樹の参加、および典型的な太平洋沿岸の存在は、オークス、チャパラル、およびバランのコミュニティを備えた茂みと森林によって増加しました。サンタモニカ市の北に位置し、サンタモニカ山脈の南端に沿って28〜26千人のサンタモニカ平野。何年も前、彼女はビルラニのコミュニティに支配されていました。サンタモニカ山脈の斜面に発達した、セットワークと犬の峡谷を備えた森林のシャパラルと孤立したパッチ。あなたが川に沿って成長した巨大なツジャとカリフォルニアの飛行機であるウィアーズビー。このような植物コミュニティは、カリフォルニア南部の海岸と同様の現代の降雨量を証言しており、その現代の範囲がさらに北に終わるセコイアの存在は、今日よりも涼しく、湿っていて季節的に変化しやすい雰囲気があったことを示しています。そのような環境は大きな草食動物を支持しました、それは獲物でした C.ドレッド そして彼の競合他社 [65] 。
獲物 [ 編集 | コードを編集します ]

生き残った動物や植物の幅広い標本、その後科学者によって抽出され検査され、過去についての知識を提供して、タールで満たされた穴に投獄されました。ランチョラブレアの穴は、南カリフォルニアのロサンゼルスの近くにあります。それらは、40万年から12、000年前に作成された粘着性のあるアスファルト堆積物のコレクションです。 400、000年前から、アスファルトはメタンによって加えられた圧力のおかげで、ギャップを通り抜けて表面に移動し、数平方メートル、深さ9〜11 mをカバーできる路を作り出します [44] 。 C.ドレッド 彼はアスファルトピットから抽出された多くの化石のために名声を得ました [70] 。主にフラグメント、2,000を超える標本がそれらから抽出されました [18] 、から生物に属します スミロドン リス、無脊椎動物、植物の場合 [44] 。ピットで表される期間には、最後の氷河最大値が含まれます。これは、世界の温度が現代よりも8°C低く、完新世の更新世の移行であり、最古のドラムを冷却します [45] [六十一] 、
同位体分析を使用して特定の化学元素を特定することができ、研究者は穴に見られる動物種の食事について結論を描くことができます。ラブレアからのスペック標本の骨コラーゲンの同位体分析は、競争の証拠を提供しました C.ドレッド 、 スミロドン 私 パンセラレオテロ 同じ獲物のために。後者は受け入れられました Camelops Hesternus 、 バイソン古い 、 Capromeryxマイナー 、 ウェストホース 私 Paramylodon Harlani 、北米大陸の草が茂った地域で放牧されています。コロンビアのマンモスと マンマットアメリカン 彼らはラブレアではまれでした。馬はさまざまな食物、フォーク – 多様な高植生を食べていましたが、最後の最大氷河と関連する植生の変化により、ラクダとバイソンは針葉樹にもっと頼らざるを得ませんでした [65] 。
化石サンプルの同位体試験 C.ドレッド 10、000年前のラブレアから、それは彼らの利益の間で馬の重要な役割の証拠を提供しましたが、あまり怠zy、マストドントン、バイソン、ラクダが食べられました [58] [66] 。これはそれを示しています C.ドレッド 彼は専門的な捕食者ではなく、彼の絶滅の前に更新世の終わりに、彼は最も利用可能な草食動物の腐肉を狩るか餌を与えました [66] 。
歯と噛む [ 編集 | コードを編集します ]


歯は口の中の歯の配置に関連しています。略奪では、上部P4施設と下部M1モルが筋肉切断ハサミや腱のように一緒に使用されるブレークマーです [71] 。
Psowateは、切断にのみ使用されるブレーカーを除き、切断と粉砕に施設を使用します。臼歯は、特定の犬の食事適応に応じて、粉砕と切断に進化した下のブレーカーを除き、粉砕に使用されます。下部ブレーカーには、層を切断するために使用されるトリゴニドがあり、粉砕に使用されるタロニドがあります。彼らの態度は、略奪的な食事の習慣を示しています。大きなトリゴニドは絶対的な肉食性を示しますが、大きなタロニドはより雑食性の食事を意味します [72] [七十三] 。変動性が低いため、下部ブレーカーの長さは略奪体のサイズを推定するために使用されます [72] 。
歯の検査はそれを示しました C.ドレッド 彼女はアメリカ大陸で最も進化的なオオカミのジャンルでした。次の機能のおかげで、他のすべてのオオカミと区別できます。 後部カスプレット P2、P3、M1メスシルイド、エントクリスト、エントコンオリド、メタコニドから超矛盾した棚に伸びる横方向のコーム、M2エントクリストおよびエントコンオニュリド上の2つのそのような結節 [74] 。
質量で制御された現代的で絶滅した略奪的な哺乳類の大規模なサンプルの牙の推定咬傷に関する研究は、体重のキログラムあたりのニュートンの胎盤の牙の咬傷が最大であることを示しました。 C.ドレッド (163)、現代の犬の中には、絶対的な肉食動物の中には、リカオンPSTREGO 142、グレイウルフ136)、ルディ112 Cyona、Dingo 108にありました。捕食者の最大の獲物は、彼の生体力学的な制限に大きく依存しています。形態 C.ドレッド 彼の現代の親relativeを思い出させました。彼がグループで狩りをした場合、現代の犬に対する彼の高い咬傷の強さは、彼が比較的大きな動物を狩ったことを示唆しています。しかし、斑点のあるロケット(117)の骨の咬傷の力は、骨の消費には牙と違反の大きな噛みつきが必要であるという広範な仮定の挑戦です。 [60] 。
頭蓋骨測定と顎の筋肉検査 C.ドレッド 15からの4つの測定された特徴を除いて、彼と現代の灰色のオオカミの間に有意な違いは見られませんでした。顎の歯は、より大きなサイズのuを除いて同じでした C.ドレッド 、P4のみが比較的大きく、より大きな刃を持ち、ブレーカーの層を切断する能力を高めました。下顎骨は比較的広く、より大きな時間的筋肉があり、灰色のオオカミよりもわずかに噛むことができました。解剖学的理由から C.ドレッド 彼は、M1およびP4の灰色のオオカミよりも頭部筋肉の小さなレバーを持っていましたが、それが機能的にどれほど重要であるかはわかりません。下部の敷地は灰色のオオカミよりも比較的わずかに大きかった [59] 、M1ははるかに大きく、切断に適していました [11] [23] [59] 。牙は、似たようなサイズの現代の犬よりも高い曲げ力を持っていました、彼らはヒエンと猫で遭遇したものに似ています [75] 。これらすべての違いは、能力を示しています C.ドレッド 灰色のオオカミよりも強い咬傷を与えるために、その柔軟で丸い牙で、彼は犠牲者との闘いに適していました [58] [59]

| 測定サイズ | 灰色のオオカミ
北米から [77] |
ラブレアのグレイウルフ [77] | ベリンギアのグレーウルフ [77] | C.と言うでしょう Zサンガモン [8] [58]
(125,000〜75,000 YBP) |
C.と言うでしょう ウィスコンシン故から [8] [58]
(50,000 YBP) |
C. Dirus Guildayi [8] [58]
(400,000〜13000年前) |
|---|---|---|---|---|---|---|
| M1の長さ | 28.2 | 28.9 | 29.6 | 36.1 | 35.2 | 33.3 |
| 幅M1 | 10.7 | 11.3 | 11.1 | 14.1 | 13.4 | 13.3 |
| トリゴニドの長さM1 | 19.6 | 21.9 | 20.9 | 24.5 | 24.0 | 24.4 |
| P4長さ | 15.4 | 16.6 | 16.5 | 16.7 | 16.0 | 19.9 |
| 幅p4 | – | – | – | 10.1 | 9.6 | 10.3 |
| P2長さ | – | – | – | 15.7 | 14.8 | 15.7 |
| P2幅 | – | – | – | 7.1 | 6.7 | 7.4 |
行動 [ 編集 | コードを編集します ]
ラ・ブレアでは、捕食性の鳥や哺乳類が死んだまたは死にかけている草食動物を惹きつけ、そこに立ち往生し、空腹の捕食者は彼らの背後にtrapに落ちました [44] [78] 。草食動物は50年ごとに彼のtrapを捕まえたと推定されています [78] そして、草食動物の遺跡の各ケースには約10の肉食動物があります [44] 。それらの最も一般的なのはラブレアです CD。ギルダイ 別 スミロドン [45] [六十一] 。残っています C.ドレッド 彼らはタールの5回の穴の灰色のオオカミの遺跡を超えています [37] 。最後の最大氷河期の間、現代のカリフォルニアの沿岸地域は、今日よりもわずかに涼しく湿った気候を持つ、彼らは避難所でした [66] 発生頻度の比較 C.ドレッド ラブレアとカリフォルニアと北米の他の地域の間の他の捕食者は、はるかに豊富であるため、数が多いことを示しています C.ドレッド ラブレア地域では、彼らはより広い地域で一般的な関係を反映していません [79] 。食べる肉食動物のほんの数だけがtrapを捕まえたと仮定することで、本当に多数のグループがあるように思われます C.ドレッド 彼らはそのような機会を一緒に食べていました [80] 。

犬の中には、性的二形性に関してわずかなばらつきがあります。テスト C.ドレッド 15360年から14310年前のTARとの穴からの日付は、頭蓋骨の長さ、牙の大きさ、下臼歯の長さに焦点を当てた穴から、オオカミで観察された灰色のオオカミと同様に、わずかな性的二形性を示しました。 C.ドレッド 一夫一婦のかさぶた [80] 。肉食動物に非常に適応した大きなサイズと高度に適応していることは、 C.ドレッド 彼は大きな獲物を食べている捕食者でした [80] [81] [82] 。フロントポーが獲物をつかむことができないため、リカオンPSTRY、Rudy Cyan、Gray Wolfは顎と下顎を使用します。これは、獲物をつかむことができないため、今年からのアルファペアとその子孫で構成されるグループで一緒に働きます。それはそれを想定することができます C.ドレッド 彼はまた、いくつかのアルファが率いる関連動物のグループに住んでいました [80] 。大規模な社会的肉食動物は、孤独な捕食者よりも大きなタールで穴に閉じ込められた獲物の死体を擁護することに成功し、グループメンバーの1人も投獄される可能性が高くなります。多くの遺体が残っています CD。ギルダイ 私 スミロドン タールと一緒に穴で見つかったことは、それが社会的捕食者についてであることを示唆しています [79] [83] 。
すべての社会的略奪哺乳類は、主に動物のメンバーの塊の合計に匹敵する体重を持つ陸上草食性哺乳類を狩ります。 [51] [84] 。かなりのサイズ C.ドレッド 彼らはあなたが300〜600 kgの成果の塊を推定することを可能にします [18] [81] [82] 。骨の安定した同位体の分析 C.ドレッド 彼女は、他の草食動物よりもバイソンなどの反min動物の好みの証拠を提供しましたが、食物の不足や太平洋沿岸沿いの海岸クジラでの炭素消費量が不足した場合に変更する可能性があります。 [18] [59] [85] 。灰色のオオカミのグループは、体重500 kgのヘラジカを倒した可能性があります。 [18] [八十六] 海岸 C.ドレッド 見事なバイソンは想像することです [18] 。いくつかの研究では、歯の骨折が噛んでスキャンダルであることを証言していることを示唆していますが、その幅広い出来事とスリムな手足は CD。ひどい 彼らは捕食者を指しています。今日の灰色のオオカミのように、 C.ドレッド 彼はおそらくブレーカーの後ろに横たわっている臼歯を使用して骨に到達し、大きいサイズにより大きな骨を分割できるようになりました [59] 。
歯の骨折 [ 編集 | コードを編集します ]
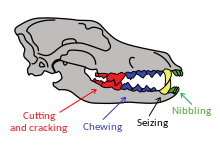
歯の骨折は、肉食動物の挙動に関連しています [八十七] 。 9つの現代の肉食動物の研究では、成人の4人に1人が歯の骨折に苦しみ、その半分が牙に関係していることが示されました。ほとんどの骨折は、斑点のある幹の幹に関係し、骨でそのすべての獲物をむさぼり食っていました。 Likaon Pstregoに最小骨折が追加されました。灰色のオオカミは真ん中にありました [八十七] [88] 。骨の食物は、比較的高い、予測不可能な緊張が明らかになっているため、ランダムな骨折のリスクを高めます。ほとんどの場合、牙は壊れ、次に小臼歯、臼歯のロッド、切歯です。フェンスを壊す最大のリスクは、その形状と役割から生じ、予測不可能な方向とサイズで曲げ方向にさらされます。大きな獲物を殺すとき、歯の骨折のリスクも高くなります [88] 。
36〜10,000のラブレア穴からの大きな捕食者の化石化された遺体の検査。数年前、それは5〜17%Uの頻度の歯の骨折を示しています C.ドレッド 、コヨーテ、アメリカンライオンと スミロドン 、現代の捕食者の0.5〜2.7%と比較して。より高い値は、すべての歯を一緒に懸念しています。牙の場合、それらは今日と違いはありませんでした。 C.ドレッド 彼は今日の灰色のオオカミよりも頻繁に切歯を壊しました。したがって、彼らは栄養中に骨の近くで働いたことが示唆されました。メキシコとペルーの化石は、同様の骨折パターンを示しています。 1993年の研究は、今日の今日よりも更新世の歯に覆われた肉食動物の骨折の頻度が高いことが、より大きな動物を狩ることから生じることがないことを示唆しています。被害者の入手可能性が低い場合、捕食者間で競争が増加し、より速く食べるために骨が沈むため、歯が骨折します。 [六十一] [八十七] [89] 。獲物が約10、000年前に消え始めたとき、それは雑食性のコヨーテを除いて、更新世の捕食者に会いました [八十七] [89] 。
La Breaの後の研究では、歯の骨折を比較しました C.ドレッド 2つの期間。穴の1つは15、000年前の化石が含まれていましたが、別の穴は13、000年でした。 15、000年前の個人は、9つの現代の肉食動物と一致する骨折が13、000年前の骨折の3倍の歯骨折で苦しんでいたことが判明しました。研究者は、15、000年から14、000年前の獲物の入手可能性が少ないか強いか、13、000年前、ゲインがホロコーストに向かっていたとき、捕食者の競争が低下し、歯の骨折の頻度もという結論に達しました。 [89] [90] 。
捕食者はグループまたは一人で狩りをすることができます。孤独なハンターは牙の噛みつきに依存しているので、強い顎が特徴です。一方、グループで狩りをしているハンターは、多くのより浅い咬傷を与えますが、顎の結合が比較的弱くなっています。したがって、研究者は化石顎の顎を使用して、狩猟方法(孤独/グループ)、さらには獲物の消費方法を決定できます。犬の顎はブレーカーの背後で強化され、動物はブレーカーの後ろに強い歯を砕いた骨(M2とM3)を砕きました。顎の強化が少なくなりました C.ドレッド グレーとレッドのオオカミよりも、しかし、コジョティとPSTREGOがリカオンに登場すると非常に似ています。背側とブリッジングエリアが悪い(P3およびP4と比較)u C.ドレッド 彼は、今日の親relativeと比較して咬傷の強さが悪いことを示しているので、グループで狩りをしなければなりませんでした。これはそれを示唆しています C.ドレッド 彼らは骨を処理することができましたが、彼らは今日の灰色のオオカミと同様に適応していませんでした [91] 。骨折の事実 C.ドレッド 今日の彼の親relativeへの後期プレイストセットの頻度の減少について [八十七] [90] 競争の減少を示唆しています C.ドレッド 骨摂取量を含む食物行動に戻るために、それはより最適でした。 [89] [91] 。その後の検査は、絶滅の直前に検出されました。現代の大きな肉食動物の中でより小さく腐肉の使用率 [92] 。
気候の影響 [ 編集 | コードを編集します ]
過去に、研究はサイズの変化が C.ドレッド それらは気候変動と相関していました [45] [六十一] [93] [九十四] [95] 。後に、それぞれ異なる時代のラブレアの4つの穴からの標本の頭蓋骨と歯の形態を比較しました。結果は、動物のサイズ、その歯と骨折、頭蓋骨と口の形状の変化を証明しています。体重 C.ドレッド それは最後の氷河の最大の最初から終わりまで落ち、暖かいアレロドゥの間にその最後に落ちました。食物欠乏の証拠は、より小さなサイズ、より大きな基部と短い口(ネオシーンの形とサイズ)の頭蓋骨、そしてより頻繁な骨折と歯の消費で現れます。 C.ドレッド 17900年の日付は、これらすべての機能を示しました。これは、食物欠乏に関連するストレスを示しています。 28、000年前の個人もこれらの特徴をある程度示していますが、それらは最大のテスト済みオオカミであったため、食物が不足していることも示唆され、オオカミはさらに大きいサイズに達しました [六十一] 。食料不足は、おそらく死体を消費し、骨を大いに壊すためにより強い咬傷をもたらしました [六十一] [96] 、そして頭蓋骨の変化により、メカニズムが改善されました。北米の気候記録は、急速な断熱材を含む氷河期の周期的な変動を明らかにし、次にDansgaard Tooschgerイベントと呼ばれる漸進的な冷却を示しています。このようなサイクルは、温度と乾燥を増加させ、ラブレアの領域で生態学的ストレスを引き起こし、それに関連して食物欠乏症を引き起こす可能性があります [六十一] 。同様の傾向が灰色のオオカミの場合に検出されました。サンタバーバラのプールでは、もともとは巨大でよく建てられており、完新世の始まりとスリムな形に置き換えられました。 [15] [26] [六十一] 。
| 特性 | 28、000年前 | 26、100年前 | 17900年前 | 13800年前 |
|---|---|---|---|---|
| 体の大きさ | 最大 | 大きい | 少しでも | 中/小 |
| 歯の骨折 | 頻繁 | レア | 頻繁 | レア |
| 歯の消費 | 大きい | 小さい | 大きい | 小さい |
| 口の形 | 短い、大きな頭蓋骨のベース | 平凡 | 短い、大きな頭蓋骨のベース | 平凡 |
| 歯の順序の形 | 信頼性のある | – | – | スリム |
| Dansgaard-oieschgerイベント | 3 | 足らない | 不正確なデータ | 不正確なデータ |
コンペ [ 編集 | コードを編集します ]

外観の少し前 C.ドレッド 北米が定住しました Xenocyon (RudyとLikaon Pstregoの祖先)、サイズ C.ドレッド そして、肉食性の大部分。化石記録はそれを珍しい動物として示しています、それは新しく形成されたもののための競争ではなかったと考えられています C.ドレッド [97] 。安定した同位体の分析は、同じ獲物の証拠を提供しました C.ドレッド 、 スミロドン そしてアメリカのライオン [65] [92] 。大きな肉食動物には、短いフットホルドベア、子鹿のプーマ、更新世の草植物コヨーテ、更新世の灰色のオオカミも含まれており、今日よりも大きくてより良いものです。これらの捕食者は、同様の獲物を探している人々と競争することができます [92] 。
ベーリングの灰色のオオカミに含まれる形態に基づいた個人は、ワイオミング州のビッグホーンバンドの基部にあるナチュラルトラップ洞窟で、25800〜14300年に放射性炭素法を使用して日付を付けられました。この場所は、ローレンティンとコルディリエルスキーの氷床の間の師団の真下にあります。氷床の間の一時的なチャネルは、これらの優れた直接的な競合他社を許可する可能性があります C.ドレッド メガファウナ狩りに適したアラスカから、南への旅のために。 C.ドレッド 彼は42°Nの北の北を実行しませんでした{後期更新世。この地域は、氷床に沿って南に広がるためにベーリングオオカミが利用できます。ベーリングオオカミがどこまで行ったかはわかりません。また、更新世後期の終わりに死亡しました [37] 。
残基 C.ドレッド それらは、北米の平原、草が茂った地域、森林地帯、南アメリカの乾燥したサバンナなど、幅広い生息地を表す地域で発見されました。これらの場所は海面から2255 mまでの高さです [17] 。遺体の配置は、これらの動物が主に開いている低地に住んでいたことと、それのための獲物を構成する大きな草食動物と一緒に住んでいたことを示唆しています [15] 。残っています C DIRE 彼らは灰色のオオカミの親relativeとは異なる重要な緯度では見られません [17] 。
アメリカの化石住民 C.ドレッド アリゾナ州、カリフォルニア州、フロリダ州、アイダホ州、インディアナ州、カンザス州、ケンタッキー州、ミズーリ州、ネブラスコ、ニューメキシコ州、オレゴン州、ペンシルベニア州、サウスカロライナ州、サウスダコタ州、テキサス州、ユタ州、バージニア州、ウエストバージニア州、ウェストバージニア州ウェストバージニア州 [17] 私はネバディ [98] 。より北の場所からの遺物のアイデンティティ [37] 。 5つのレポートは、未確認の遺跡について語っています C.ドレッド オレゴン州の化石湖の北42度(125000年から10000年以前)、アイダホ州のアメリカ滝貯水池(125000〜75000年)、南ダコタ州のサンマンダー洞窟(2、500,000年)、北のネプレイズの4つの近い場所(250,000年前の場所) [37] )。これは、温度、獲物、生息地のために種の範囲を制限することを示唆しています [37] 。発見を提供する主な場所 CD。ひどい 彼らはロッキー山脈の東にあり、テキサスのサンアントニオ近くのフリーゼンハーンの洞窟を覆っています。 [18] 。
中央および南東環境のメキシコの10の場所は、このジャンルの遺跡で知られています。カリフォルニア州バハのコモンド市。サンルイスポトシのエルセドラル。連邦地区のプエブラ市の近くのテキクスキーアトとバルケリーのエルタホ採石場。ハリスコのラゴ・デ・チャパラ。ユカタンのloltun洞窟。シナロアのポトラシトとヌエボレオンのアランベリ近くのサンジョセシト洞窟洞窟。中央メキシコの場所から、サンホセシトとエルセダゾは最大の個人を提供しました C.ドレッド ある町から。他の場所ではわずかな標本のみが見つかりました。ソノラの検査が確認されました CD。ギルダイ [58] 。
南アメリカで C.ドレッド 日付は17,000年未満にさかのぼります。彼らは3つの場所でのみ発見されました:ムアコ、ベネズエラのファルコン。ペルーのタララ州とボリビアのタリヤ局 [17] 。もしも C.ドレッド それは北米で進化し、実際にはアンデスの廊下を通して南アメリカで広がっています [17] [22] 、中程度の哺乳類が中央アメリカから南アメリカに移動した提案された道は、当時のこれらの地域を特徴付ける涼しく乾燥したオープンな生息地のために移動しました。ほとんどの場合、これは氷河の間に開催されました。なぜなら、廊下はオープンで乾燥したエリアとサバンナで構成されていたため、Inlglacilaでは、熱帯熱帯雨林が彼と一緒に生い茂っていたため [17] [99] 。
描画は、種の地理的除去の結果であり、人口の減少はゼロまで減少します。生物地理学的範囲と人口に影響を与える要因には、競争、捕食者の相互作用、物理的環境の変化、ランダムイベントが含まれます [100] 。アメリカのメガファウナは12.7千人で亡くなりました数年前。次に、質量が44 kgを超える90種類の哺乳類が絶滅しました [45] [六十一] 。絶滅の提案されたモデルの1つ [17] [八十七] [101] [102] 。また、メガロを食べる人の絶滅が絶滅を説明することも示唆されています C.ドレッド 両方のアメリカ大陸で [17] 。 2007年の調査は、ヨーロッパ、シベリア、アメリカの環境条件に焦点を当てていました。 10,000数年前。それは、脱直につながる長期にわたる断熱材と、草食動物に耐える植生で覆われた広大な湿地への大きな草食動物の高草食動物の変換の直前に現れる最高の降雨量を示しました。湿気に依存する環境の変化がメガファウナの絶滅につながったことが提案されています。そのため、アフリカの祭までの場所が彼女の牧草地を救い、砂漠と森林の間にさらに存在し、それゆえそこにあまり偉大な種を描きませんでした [103] 。
ラブレアのラスイーター種の歯のエナメル質のマイクロインジュリーの結果、 C.ドレッド 、肉食動物は、絶滅の直前に食物欠乏に関連するストレスで脅かされなかったことを示唆しています。さらに、腐肉の利用(これは、骨折や骨を食べることを含む必要な量の腐肉の消費)でした)は、今日よりも大きな肉食動物の間でした [92] 。別の研究では、生物地理学についてはほとんど知られていないため、絶滅のモデルをいくつか調べなければならないことが示唆されました。 C.ドレッド 、その潜在的な競合他社と成果、またはこれらの種が絶滅中の環境の変化にどのように影響したか、またはどのように反応したか [17] 。絶滅 C.ドレッド それは、ミアロを食べる人、気候変動の影響、人々を含む他の種との競争に依存することに基づいている可能性があります [92] しかし、その理由は論争の対象のままです [104] 。最年少の遺跡は、ミズーリ州のブーンのブリンジュルソン湾で発見されました。彼らは9440年にさかのぼります [23] 。
- ↑ a b J.ライディ。 ニオブララ川の谷からの絶滅した脊椎の遺物の通知は、1857年の探検中にネブラスカで、リュートの指揮の下で集められました。 G. K.ウォーレン、米国トップ。 Eng。、F。V.ヘイデン博士、地質学者への遠征隊、議事録 。 「フィラデルフィアの自然科学アカデミー」。 10、s。 21、1858。
- ↑ a b J.ライディ。 インディアナ州オハイオ川のほとりにあるフランシス・A・リンクス氏によって発見されたいくつかの化石骨の通知 。 „議事録:フィラデルフィアの自然科学アカデミー」、s。 200、1854。
- ↑ a b J.ライディ。 ダコタとネブラスカの絶滅した哺乳類の動物相は、他の地域からのいくつかの連合形態の説明を含む、北米の哺乳類の遺跡の概要とともに 。 「フィラデルフィアの自然科学アカデミーのジャーナル」。 7、s。 368、1869。
- ↑ a b はい。全て。 アッパーミシシッピ州の鉛地域からのオオカミの絶滅した種と絶滅した鹿のいくつかの残骸の説明 。 「アメリカンジャーナルオブサイエンス」。 S3-11(61)、s。 47–51、1876。doi: 10.2475/ajs.s3-11.61.47 。
- ↑ E.H.セラーズ。 フロリダの更新世からの人間の遺体と関連する化石 。 「フロリダ地質調査所の年次報告書」。 8、s。 152、1916。
- ↑ a b J.C.メリアム。 カニス・ディルス・グループのオオカミの体系的な位置に注意 。 「カリフォルニア大学地質学部の紀要」。 10、s。 533、1918。
- ↑ a b C.フリック。 アラスカの冷凍動物相 。 「自然史」、s。 71–80、1930。
- ↑ a b c d そうです f g h B. Kurten:北米のRancholabrean Dire Wolf(Canis Dirus Leidy)の地理的違い。 W:H.H。ジェノウェイズ、M.R。ドーソン: 第四紀脊椎動物の古生物学の貢献:ジョン・E・ギルデイの記念巻 。カーネギー自然史博物館、1984年、s。 218 –227、Seria:Special Publication 8. ISBN 978-0-935868-07-4 。
- ↑ ページ 美術館 、 Rancho La Breaでコレクションをご覧ください 、[w:]ロサンゼルス郡財団の自然史博物館[オンライン] [アクセス2017-10-01] [アーカイブ 住所 2017-01-25] 。
- ↑ テント 、 オンラインコード(第6章、第23.1条を参照) 、[W:]国際動物学の命名法のオンライン[オンライン]、動物学的命名法の国際委員会、2017年 [アクセス2017-06-09] [アーカイブ 住所 2009-05-24] 。
- ↑ a b c d そうです f g J.C.メリアム。 ランチョ・ラ・ブレアの動物相、パートII。カニダエ。 。 「カリフォルニア大学の回顧録」。 1、s。 217–273、1912。
- ↑ E.L.トロクセル: テキサス州ロッククリークの脊椎動物の化石 。 T. 189. American Journal of Science、1915、s。 613–618。
- ↑ マークスティーブンソン:9。W:ロバータL.ホール、ヘンリーS.シャープ: オオカミと男:並行して進化 。ニューヨーク:Academic Press Inc、1978、s。 180. ISBN 978-0-12-319250-9 。 [アクセス2017-10-01]。
- ↑ E.L.ルンデリウス。 化石脊椎動物、テキサス州サンパトリシオ郡、更新世後期のイングレスサイド動物相 。 「経済地質局」。調査No.77の報告、s。 1–74、1972。
- ↑ a b c d Nowak 1979↓ 。
- ↑ X.ワン。 カンザス川には、カンザス州の悲惨なオオカミに関するメモがある更新世の悲惨なオオカミが残っています 。 「カンザス大学自然史博物館の時折論文」。 137、s。 1–7、1990。
- ↑ a b c d そうです f g h 私 j k l m n o R.G.ダンダス。 悲惨なオオカミの第四紀の記録、 犬の恐怖 、北アメリカと南アメリカで 。 「ボレアス」。 28(3)、s。 375–385、1999。doi: 10.1111/j.1502-3885.1999.tb00227.x 。
- ↑ a b c d そうです f g h 私 j k ウィリアム・アニョンゲ;クリス・ローマン。 の新しい体重推定 カニス Dirus、絶滅した更新世の悲惨なオオカミ 。 「脊椎動物の古生物学のジャーナル」。 26、s。 209–212、2006。doi: 10.1671/0272-4634(2006)26 [209:nbmefc] 2.0.co; 2 。
- ↑ 2008年のテッドフォードの王。↓ 、s。 148。
- ↑ a b c テッドフォード、ワン・アイ・テイラー2009↓ 、s。 181。
- ↑ R.A.マーティン; S. D.ウェッブ:レヴィ郡の悪魔のデン・ファウナの更新世後期哺乳類。 W:S.D。ウェッブ: フロリダの更新世哺乳類 。ゲインズビル:フロリダ大学出版局、1974年、s。 114 –145。 ISBN 978-0-8130-0361-0 。
- ↑ a b Berta 1988↓ 。
- ↑ a b c d 11カルマー味。 W:B。カーテン; E.アンダーソン: 北米の更新世哺乳類 。コロンビア大学出版局、ニューヨーク、1980年、s。 168–172。 ISBN 978-0-231-03733-4 。
- ↑ ヴァレリウススピリット: 世界の鹿:彼らの進化、行動、エコロジー 。 WYD。 1. Stackpole Books、Mechanicsburg、Pennsylvania、1998、s。 10. ISBN 0-8117-0496-3 。 [アクセス2017-05-01]。
- ↑ ヴァレリウススピリット。 氷河期哺乳類の種分化について、特に頸部とカプリッドを参照して 。 「カナダズロジージャーナル」。 65(5)、s。 1067、1987。doi: 10.1139/Z87-171 。
- ↑ a b G.D. Goulet: 近距離、現代、完新世、および後期更新世の灰色オオカミ(カニスループス)と選択されたカニス(修士論文)の間の側頭および地理的スカルの変動の比較 。マニトバ大学、ウィニペグ、1993年、s。 1–116。
- ↑ a b 2008年のテッドフォードの王。↓ 、s。 52。
- ↑ ロナルド・M・ノバック;ニコラスE.(ブルスコ)フェデラル。 イタリアのオオカミの体系的なステータス カニス ループス 。 「Acta Theriologica」。 47(3)、s。 333–338、2002。doi: 10.1007/BF03194151 。
- ↑ Nowak 2003↓ 、s。 243。
- ↑ a b c d テッドフォード、ワン・アイ・テイラー2009↓ 、s。 146。
- ↑ a b テッドフォード、ワン・アイ・テイラー2009↓ 、s。 144。
- ↑ Nowak 2003↓ 、s。 242。
- ↑ a b Prevosti 2010↓ 、s。 472。
- ↑ Nowak 1979↓ 、s。 93。
- ↑ E.L. Lundelius、C.J。Bell:7 Blancan、Irvingtonian、およびRancholabrean Mammal Ages。 W:マイケルO.ウッドバーン: 北米の白亜紀後期および新生代の哺乳類:生物層序と地質学 。コロンビア大学出版局、2004年、s。 285. ISBN 978-0-231-13040-0 。 [アクセス2017-05-01]。
- ↑ a b J. I.ミード; C.マンガナロ; C. A.リポーション; L. D. Agenbroad:サウスダコタ州ブラックヒルズのサラマンダー洞窟の初期のrancholabrean哺乳類。 W:K.M。スチュワート、K.L。シーモア: 新生代後期哺乳類の古生物学と古環境、C.S。(ルーファス)のキャリアへの敬意 。トロント大学出版局、カナダ、オンタリオ州トロント、1996年、s。 458–482。 ISBN 978-0-8020-0728-5 。
- ↑ a b c d そうです f Meachen、Brannick I Fry 2016↓ 。
- ↑ a b テッドフォード、ワン・アイ・テイラー2009↓ 、s。 148。
- ↑ Berta 1988↓ 、s。 113。
- ↑ Nowak 1979↓ 、s。 116。
- ↑ a b アンジェラR. ペリ 私はイニ 、 悲惨なオオカミは古代の新世界の缶の系統の最後でした 、「自然」、2021、doi: 10.1038/s41586-020-03082-x 、PMID: 33442059 。
- ↑ D.N. Janczewski 私はイニ 、 ランチョラブレアのセイバー歯の猫化石からの分子系統発生推論 、「アメリカ合衆国科学アカデミーの議事録」、89(20)、1992年、s。 9769–9773、doi: 10.1073/pnas.89.20.9769 、PMID: 1409696 、PMCID: PMC50214 。
- ↑ 命を与える。 金 私はイニ 、 Rancho La Brea Columbian Mammoth(Mammuthus Columbi)からのDNA抽出の試み:アスファルト堆積物からの古代DNAの見通し 、「生態学と進化」、4(4)、2014年、s。 329–336、doi: 10.1002/ECE3.928 、PMID: 24634719 、PMCID: PMC3936381 。
- ↑ a b c d そうです C.株:株式: ランチョ・ラ・ブレア:カリフォルニアでの更新世の生活の記録 。 WYD。 7.ロサンゼルス郡自然史博物館、1992年、s。 1–113、Seria:Scienceシリーズ。 ISBN 978-0-938644-30-9 。
- ↑ a b c d そうです F.R.オキーフ; E.V. fet; J.M.ハリス。 動物相と花の放射性炭素の日付の編集、キャリブレーション、および合成、カリフォルニア州ランチョ・ラ・ブレア 。 「科学への貢献」。 518、s。 1–16、2009。
- ↑ Meachen、Brannick I Fry 2016↓ s。 T1。
- ↑ a b テッドフォード、ワン・アイ・テイラー2009↓ 。
- ↑ a b Mech 1966↓ 、s。 1。
- ↑ a b c d Adam Hartstone-Rose;ロバート・G・ダンダス;ブリッティン・ボイド;ライアンC.ロング; Aisling B. Farrell;クリストファー・A・ショー:ジョン・M・ハリス: ランチョ・ラ・ブレアのバキュラ 。 T.科学における貢献(ラブレアとそれ以降の特別なボリューム:アスファルト保存されたビオタの古生物学は、ロサンゼルス郡のランチョラブレアでのロサンゼルス郡の発掘博物館の100周年を記念して)を記念して)。自然史博物館ロサンゼルス郡、2015年、s。 53–63、SERIA:科学シリーズ42。
- ↑ a b c d チェスター株;ジョン・F・ランス。 カニス・ディルスの四肢要素の相対長 。 「南カリフォルニア科学アカデミーの紀要」。 47(3)、s。 79–84、1948。
- ↑ a b ボリス・ソーキン。 陸生哺乳類の捕食者の体重に対する生体力学的制約 。 「レダイア」。 41(4)、s。 333-347、2008。doi: 10.1111/j.1502-3931.2007.00091.x 。
- ↑ グレイウルフ(ユーコンの) 、[W:]環境ユーコン[オンライン]、カナダ政府、2017年 [アクセス2017-04-18] 。
- ↑ ランチョ・ラ・ブレア。チャス。 R.ナイト。壁画amer。博物館の殿堂入り。背景の海岸の範囲、古いbaldy、 [初め] 。
- ↑ Theodosius Dobzhansky:ダーウィン生物学の基本的な概念について。 W:Theodosius Dobzhansky、Max K. Hecht、William C. Steere: 進化生物学 。 T. 2.ニューヨーク:Appleton-Century-Crofts、1968、s。 1–34。 doi: 10,1007/978-1-4684-8094-8_1 。 OCLC 24875357 。
- ↑ アンジェラペリ。 犬の衣服のオオカミ:最初の犬の家畜化と更新世のオオカミのバリエーション 。 「考古学科学のジャーナル」。 68、s。 1–4、2016。doi: 10.1016/j.jas。2016.02.003 。
- ↑ レナード、ジェニファー。 エコロジーは、灰色のオオカミの進化を促進します 。 「進化生態研究」。 16、s。 461–473、2014。
- ↑ ルーシーO.H.花;ダニエルC.シュリーブ。 ヨーロッパの更新世のカニドにおける古遠足の変動の調査 。 「第四紀科学のレビュー」。 96、s。 188–203、2014。doi: 10.1016/j.quascirev.2014.04.015 。 bibcode: 2014QSRV … 96..188F 。
- ↑ a b c d そうです f g ジョン・ポール・M・ホドネット、ジム・I・ミード、A。バエズ。 オオカミと言う、 犬の恐怖 (哺乳類、Carnivora; Canidae)、メキシコの東中央ソノラの更新世後期(rancholabrean)から 。 「南西の自然主義者」。 54.1、s。 74–81、2009。doi: 10.1894/clg-12.1 。
- ↑ a b c d そうです f W. Santyonge; A.ベイカー。 Canis Dirusの頭蓋顔面の形態と摂食行動、絶滅した更新世の悲惨なオオカミ 。 「Zoologyのジャーナル」。 269(3)、s。 309–316、2006。doi: 10.1111/j.1469-7998.2006.00043.x 。
- ↑ a b S. wroe; C. MCHENRY; J.トーマソン。 バイトクラブ:大きな噛む哺乳類の比較咬合力と化石分類群における略奪行動の予測 。 「王立協会の議事録B:生物科学」。 272(1563)、s。 619–625、2005。Doi: 10.1098/rspb.2004.2986 。 PMID: 15817436 。 PMCID: PMC1564077 。
- ↑ a b c d そうです f g h 私 j k F.ロビンオキーフ;ウェンディJ.バインダー;スティーブン・R・フロスト;ルドヤードW.サドリエ;ブレアヴァンバルケンバーグ。 Rancho La BreaのDire Wolf、Canis Dirusの頭蓋形態計測:時間的変動性と栄養ストレスと気候へのリンク 。 「Palaeontologia Electronica」。 17(1)、s。 1–24、2014。
- ↑ a b 気候変動に関する政府間パネル(UN) 、 IPCC 4番目の評価レポート:気候変動2007 – 古気象観点 、ノーベル財団、2007年 [アクセス2017-06-09] [アーカイブ 住所 2018-07-22] 。
- ↑ a b P. U.クラーク; A. S.ダイク; J.D. Shakun; A. E.カールソン; J.クラーク; B. Wohlfarth; J. X. Mitrovica; S. W. Hostetler; A. M.マッケイブ。 最後の氷河最大 。 “化学”。 325(5941)、s。 710–714、2009。doi: 10.1126/science.1172873 。 PMID: 19661421 。 bibcode: 2009Sci … 325..710C 。
- ↑ S.A. エリアス 、 D. 書かれています 、 更新世後期の巨大な絶滅 、「地球システムと環境科学の参照モジュール」、2016年、s。 3202–3217、doi: 10.1016/B978-0-12-409548-9.10283-0 、ISBN 978-0-12-409548-9 。
- ↑ a b c d ジョーン・ブレンナー・コルトレイン。ジョン・M・ハリス; Thure E. Cerling;ジェームズ・R・エーレリンガー;マリア・デニーズ・ディアリング;ジョイウォード;ジュリー・アレン。 Rancho la brea安定同位体生物地球化学と、南カリフォルニア沿岸沿岸の更新世後期の古生態学に対するその意味 。 「古地理学、古気候学、古生態学」。 205(3–4)、s。 199–219、2004。doi: 10.1016/j.palaeo.2003.12.008 。
- ↑ a b c d K. Fox-Dobbs; J.K.バンプ; R.O.ピーターソン; D.L.狐; P.L.コッホ。 肉食動物固有の安定性同位体変数と現代および古代のオオカミの集団の採餌生態学の変動:ミネソタ州アイル・ロワイヤルのケーススタディ 。 「カナダズロジージャーナル」。 85(4)、s。 458–471、2007。doi: 10.1139/Z07-018 。 [アクセス2017-05-06]。
- ↑ マイケルJ.モラート: カリフォルニアの考古学 。アカデミックプレス、フロリダ州オーランド、1984年、s。 89. ISBN 0-12-506180-3 。 [アクセス2017-05-01]。
- ↑ ドナルド・リー・ジョンソン。 カリフォルニア沿岸後期の第四紀の気候:氷河期避難所の証拠 。 「第四紀研究」。 8(2)、s。 154、1977。Doi: 10.1016/0033-5894(77)90043-6 。 bibcode: 1977Qures … 8..154J 。
- ↑ 芸術大学と 科学 、 ランチョラブレアタールプール。 W.B.のブルースホースフォールによる修復スコット 、[W:]ケースウエスタンリザーブ大学[オンライン]、2016年 [アクセス2016-12-24] 。
- ↑ 2008年のテッドフォードの王。↓ 、s。 149。
- ↑ 2008年のテッドフォードの王。↓ 、s。 74。
- ↑ a b ガブリエレサンサロン; Davide FedericoBertè;レオナルド・マオリノ;ルカ・パンドルフィ。 ヨーロッパの更新世のオオカミカニスループス(哺乳類、カニダエ科)のカルナシアル歯の進化的傾向と停滞 。 「第四紀科学のレビュー」。 110、s。 36、2015。doi: 10.1016/j.quascirev.2014.12.009 。
- ↑
 マルコ・チェリン; Davide FedericoBertè; Raffaele Sardella;ロレンツォルーク。 Canis Etruscus(Canidae、Mammalia)とPantalla(Perugia、Central Italy)からの動物相の集合におけるその役割:イタリアの故ヴィラフランチアの大肉食性ギルドとの比較 。 「イタリアの古生物学会の速報」。 52(1)、s。 11–18、2013。[Dostęp2018-03-26]。
マルコ・チェリン; Davide FedericoBertè; Raffaele Sardella;ロレンツォルーク。 Canis Etruscus(Canidae、Mammalia)とPantalla(Perugia、Central Italy)からの動物相の集合におけるその役割:イタリアの故ヴィラフランチアの大肉食性ギルドとの比較 。 「イタリアの古生物学会の速報」。 52(1)、s。 11–18、2013。[Dostęp2018-03-26]。 - ↑ Berta 1988↓ 、s。 50。
- ↑ B. Van Valkenburgh; C. B.ラフ。 大きな肉食動物の犬の歯の強さと殺害行動 。 「Zoologyのジャーナル」。 212(3)、s。 379–397、1987。doi: 10.1111/j.1469-7998.1987.tb02910.x 。
- ↑ J.C. メリアム 、 ランチョ・ラ・ブレアの動物相 、t。 1、カリフォルニア大学 – バークレー、1911年、s。 224–225 。
- ↑ a b c ジェニファーA. レナード 私はイニ 、 メガファウナールの絶滅と専門のオオカミのecomorphの消失 、「現在の生物学」、13、17、2007、s。 1146–1150、doi: 10.1016 / j.cub.2007.05.072 、PMID: 17583509 [アクセス2017-05-06] 。
- ↑ a b L. F.マーカス; R.バーガー:ランチョラブレアの放射性炭素日付の重要性。 W:P.S。マーティン、R.G。クライン: 第四紀絶滅 。アリゾナ大学出版局、ツーソン、1984年、s。 159–188。 ISBN 978-0-8165-0812-9 。
- ↑ a b ブリアナ・K・マコーゼ;ジョン・D・オーカット;エドワード・B・デイビス。 Rancho LabaのCarnivoran Fauna:平均または異常? 。 「古地理学、古気候学、古生態学」。 329–330、s。 118–123、2012。doi: 10.1016/j.palaeo.2012.02.022 。
- ↑ a b c d ブレアヴァンバルケンバーグ。タイソン・サッコ。 大規模な更新世の肉食動物における性的二形性、社会的行動、および概念内競争 。 「脊椎動物の古生物学のジャーナル」。 22、s。 164–169、2002。doi: [0164:Sdsbai 2.0.co; 2 10.1671/0272-4634(2002)022 [0164:SDSBAI] 2.0.co; 2]。
- ↑ a b B. Van Valkenburgh; K.-P. Koepfli:Canidsの捕食への頭蓋および歯の適応。 W:N。ダンストン、J.L。ゴーマン: 捕食者としての哺乳類 。オックスフォード大学出版局、オックスフォード、1993年、s。 15–37。
- ↑ a b B. Van Valkenburgh:更新世後期の北米の捕食者の衰退。 W:J.J。サンダース、B.W。スタイル、G.F。バリシニコフ: 北半球における第四紀の古zoology 。イリノイ州立博物館の科学論文、スプリングフィールド、1998年、s。 357–374。 ISBN 978-0-89792-156-5 。
- ↑ C. 炭素 私はイニ 、 再生と更新世のタールの浸透の類似点は、絶滅したセイブトゥース猫の社会性を示唆しています、smilodon 、「生物学の文字」、5(1)、2009年、s。 81–85、doi: 10.1098/rsbl.2008.0526 、PMID: 18957359 、PMCID: PMC2657756 。
- ↑ M.アール。 社会的肉食動物の柔軟な体重 。 「アメリカの自然主義者」。 129(5)、s。 755–760、1987。doi: 10.1086/284670 。
- ↑ K. Fox-Dobbs; P. L. Koch; M. T.クレメンツ。 Labreaでの昼食:Smilodon FatalisとCanis Dirusの食事パターンの等型再構築時間 。 「脊椎動物の古生物学のジャーナル」。 23(3、補足)、s。 51a、2003年。Doi: 10,1080/02724634.2003.10010538 。
- ↑ Mech 1966↓ 、p。76、そうすることができます。
- ↑ a b c d そうです f ブレアヴァンバルケンバーグ。フリッツヘルテル。 ラブレアでの厳しい時代:更新世後期の大きな肉食動物の歯の切断 。 「科学、新シリーズ」。 261(5120)、s。 456–459、1993。アメリカ科学進歩協会。 doi: 10.1126/science.261.5120.456 。 PMID: 17770024 。 bibcode: 1993sci … 261..456V 。
- ↑ a b B Van Valkenburgh。 大規模な略奪的な哺乳類における歯の切断の発生率 。 “私は持っている。 nat。」。 131(2)、s。291–302、1988。 10.1086/284790 。
- ↑ a b c d ブレアヴァンバルケンバーグ。 肉食性のコスト:更新世と最近の肉食者の歯の骨折 。 「リンネ協会の生物学的ジャーナル」。 96、s。 68–81、2008。doi: 10.1111/j.1095-8312.2008.01108.x 。
- ↑ a b ウェンディJ.バインダー;エリシア・N・トンプソン;ブレアヴァンバルケンバーグ。 ランチョラブレアダイアオオカミの間の歯の骨折の時間的変動 。 「脊椎動物の古生物学のジャーナル」。 22(2)、s。 423–428、2002。doi: 10.1671/0272-4634(2002)022 [0423:TVITFA] 2.0.co; 2 。
- ↑ a b フランソワ・テリエン。 現存する肉食動物の下顎力プロファイルと絶滅した捕食者の摂食挙動への影響 。 「Zoologyのジャーナル」。 267(3)、s。 249、2005。Doi: 10.1017/s0952836905007430 。
- ↑ a b c d そうです L.R.G.デサンティス; B.W.シューベルト; E.シュミットリンビル; P. ungar; S.ドノヒュー; R.J.ハウプト:ジョン・M・ハリス: カリフォルニア州ラブレアタールピットからの肉食動物の歯科用マイクロウェアテクスチャと潜在的な絶滅の意味 。 T.科学における貢献(ラブレアとそれ以降の特別なボリューム:アスファルト保存されたビオタの古生物学は、ロサンゼルス郡のランチョラブレアでのロサンゼルス郡の発掘博物館の100周年を記念して)を記念して)。ロサンゼルス郡自然史博物館、2015-08-15、s。 37–52、Seria:Scienceシリーズ42。
- ↑ J.O. nigra; J.F.ランス。 Dire Wolf Groupのメタポディアルの統計研究 。 「南部科学アカデミーの紀要」。 46、s。 26–34、1947。
- ↑ C.A.ショー; A.テハダフロア。 Rancho La BreaのSmilodon ectocuneiformsの変動の生体力学的意味 。 「科学における貢献」。 359、s。 1–8、1985。
- ↑ F.R.オキーフ。 上部更新世の気候変動に対する悲惨なオオカミの人口レベルの反応、カニス・ディルス 。 「脊椎動物の古生物学のジャーナル」。 28、s。 122a、2008。doi: 10,1080/02724634.2008.10010459 。
- ↑ Zhijie Jack Tseng 、 Xiaoming 王 、 化石犬の頭蓋機能形態と、ボロファガスとエピシオンのdurophagyのための適応(Carnivora、Mammalia) 、「Journal of Morphology」、11、271、2010、s。 1386–98、doi: 10.1002/jmor.10881 、PMID: 20799339 。
- ↑ 2008年のテッドフォードの王。↓ 、s。 60。
- ↑ エリック・スコット;キャスリーンB.スプリンガー。 の最初のレコード カニス dirusと スミロドン ネバダ州アッパーラスベガスウォッシュの地元の動物相は、更新世後期のツールスプリングスからのファタリス 。 「Peerj」。 4、p。E2151、2016。Doi: 10.7717/peerj.2151 。 PMID: 27366649 。 PMCID: PMC4924133 。
- ↑ SD。ウェッブ: 生態地理学と偉大なアメリカのインターチェンジ 。 WYD。 3. T. 17. Paleobiology、1991、s。 266–280。 doi: 10.1017/s0094837300010605 。
- ↑ スティーブン・M・スタンリー: 絶滅 。 Scientific American Library、ニューヨーク、1987年、s。 242。
- ↑ R. W.グラハム; J. I.ミード:北米での最後の脱膜中の哺乳類動物相の環境変動と進化。 W: 最後の脱副化中の北米と隣接する海洋 。 Geological Society of America K-3、ボルダー、コロラド、1987年、s。 371–402。 ISBN 978-0-8137-5203-7 。 著者を確認してください:1と3。
- ↑ A. D. Barnosky:広範囲にわたる哺乳類の絶滅のパラダイムとしての更新世後期絶滅イベント。 W:スティーブンK.ドノバン: 大量絶滅:プロセスと証拠 。コロンビア大学出版局、ニューヨーク、1989年、s。 235–255。 ISBN 978-0-231-07091-1 。
- ↑ M. Timothy Rabanus-Wallace;マシュー・J・ウーラー;グラントD.ザズラ;エレン・シュート; A.ホープジャーレン。 Pavel Kosintsev;ジェームズ・A・バーンズ;ジェームズ・ブリーン;バスティエン・ラマス;アランクーパー。 メガファウナル同位体は、更新世後期の絶滅中に放牧地での水分の増加の役割を明らかにしています 。 「自然のエコロジーと進化」。 1(5)、s。 0125、2017。doi: 10.1038/s41559-017-0125 。
- ↑ アレクサンドリア・L・ブラニック;ジュリーA.ミーチェン; F.ロビン・オキーフ:ジョン・M・ハリス: ランチョ・ラ・ブレアの悲惨なオオカミ、カニス・ディルスの顎の形の微小進化 。 T.科学における貢献(ラブレアとそれ以降の特別なボリューム:アスファルト保存されたビオタの古生物学は、ロサンゼルス郡のランチョラブレアでのロサンゼルス郡の発掘博物館の100周年を記念して)を記念して)。自然史博物館ロサンゼルス郡、2015年、s。 23–32、SERIA:科学シリーズ42。
![[初め]](http://geology.cwru.edu/~huwig/catalog/slides/748.B.17.jpg){kind=link}
{kind=link}
- A. ベルタ 、 南アメリカの大規模なカニダエ科の第四紀の進化と生物地理学(哺乳類:カルニボラ) 、「地質科学におけるカリフォルニア大学出版物」(132)、1988年、s。 1–49、ISBN 978-0-0-09-0960-9 。
- ジュリーA. ミーチェン 、 アレクサンドリアL. ブラニック 、 トレントJ. 稚魚 、 米国大陸で見られる絶滅したベリンギャンのオオカミの形態型は、オオカミの移住と進化に影響を及ぼします 、「生態学と進化」、6(10)、2016年、s。 3430、doi: 10.1002/ECE3.2141 、PMID: 27252837 、PMCID: PMC4870223 。
- L.デイビッド メカ 、 アイル・ロワイヤルのオオカミ 、1966年の米国国立公園の動物相(動物相シリーズ7)、ISBN 1-4102-0249-6 [アクセス2017-05-01] 。
- ロナルドM. Nowak 、 北米の第四紀カニス 、カンザス大学自然史博物館のモノグラフ、1979年、s。 106、doi: 10.5962/bhl.title.4072 、ISBN 978-0-89338-007-6 [アクセス2017-05-01] 。
- R.M. Nowak 、 9ウルフの進化と分類 、 [の:] L.デイビッド メカ 、 ルイージ ビチャニ (赤。)、 オオカミ:行動、生態学、保全 、シカゴ大学出版局、2003年、ISBN 978-0-26-51696-7 。
- フランシスコJ. 前世代 、 「完全な証拠」アプローチを使用して、南アメリカの大規模な南アメリカの缶詰(哺乳類、carnivora、canidae)の系統発生 、「cladistics」、26(5)、2010、s。 456–481、doi: 10.1111/j.1096-0031.2009.00298.x 。
- リチャードH. テッドフォード 、 Xiaoming 王 、 ベリルE. テイラー 、 北米の化石カニナエの系統系統系(Carnivora:Canidae) 、「アメリカ自然史博物館の紀要」、325、2009、s。 1–218、doi: 10,1206/574.1 。
- Xiaoming 王 、 リチャードH. テッドフォード 、 犬:化石の親relativeと進化の歴史 、コロンビア大学出版局、ニューヨーク、2008年、s。 1–232、ISBN 978-0-231-13529-0 。
Recent Comments