Acide γ-aminobutyryque-Wikipedia
Homonimiczne artykuły patrz GABA.
| Kwas γ-aminobutyryque | ||
 |
||
 |
||
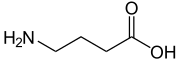
| Struktura GABA. | ||
| Identyfikacja | ||
|---|---|---|
| Nazwa UICPA | Kwas 4-aminobutanowy | |
| N O Cas | ||
| N O Echa | 100 000,235 | |
| N O TEN | 200-258-6 | |
| Pubchem | 119 | |
| FEMA | 4288 | |
| Chemikalia | ||
| Formuła | C 4 H 9 NIE 2 [Izomery] | |
| Masa cząsteczkowa [[[ Pierwszy ] | 103 119 8 ± 0,004 6 g/mol C 46 59%, H 8,8%, N 13 58%, o 31 03%, |
|
| P K A | 40459 | |
| Właściwości fizyczne | ||
| T ° połączenie | 203 ° C. (rozkład) [[[ Ref. pożądany] | |
| Rozpuszczalność | 1 300 G · L -Pierwszy w wodzie przy 25 ° C. [[[ Ref. pożądany] | |
| Środki ostrożności | ||
| Dyrektywa 67/548/EEC | ||
 Xi |
||
| Ecotoksykologia | ||
| Dl 50 | 12 680 Mg · kg -Pierwszy (Mysz, Perral) [[[ Ref. pożądany] | |
|
|
||
| To jednostki i Cntp , o ile nie zaznaczono inaczej. | ||
| modyfikator |
||
L ‘ kwas γ-aminobutyryque , często skrócone w PRZÓD (z angielskiego G Ale- A Uważać B Utyric A CID ), jest głównym inhibitorem neuroprzekaźnika ośrodkowego układu nerwowego u ssaków i ptaków. U owadów jest obecny w całym ciele. Jest to neuromodulator [[[ 2 ] Uznany za hamujący u dorosłych, ale ekscytujący podczas rozwoju embrionalnego człowieka. Odgrywa ważną rolę u dorosłych, zapobiegając długotrwałym wzbudzeniu z neuronów. Ma również rolę neurotroficzną, to znaczy, że promuje wzrost niektórych neuronów.
Hamujące działanie GABA przeciwwagi wzbudzające działanie glutaminianu. Nierównowaga między tymi dwoma neuroprzekaźnikami jest zaangażowana w padaczkę i niedokrwienie mózgu. Jego wzór brutto to c 4 H 9 N O 2 .
GABA byłby zaangażowany w co najmniej 30% [[[ 3 ] Synapsy mózgu. Drugim hamującym neuroprzekaźnikiem jest glicyna, która jest szczególnie zlokalizowana w szpiku kręgosłupa.
Dwa rodzaje neuronów gabacinergicznych spotykają się:
GABA wykryto głównie w neuronach nieramidalnych, takich jak komórki ziarniste lub gwiazdorskie, w warstwach II do IV kory. Postulowano, że interneurony gabacinergiczne wywierają modulacyjny wpływ na ciało komórki i początkowy segment aksonów komórek piramidalnych (glutaminergiczny), aby utrzymać trwałą aktywność w utrzymaniu pamięci pamięci roboczej [[[ 6 ] .
GABA jest syntetyzowany z kwasu glutaminowego przez enzym GD ( Glutaminian dékarboksylaza ) Lub AGD ( Kwas glutaminowy ) [Gad w języku angielskim ( Dekarboksylaza kwasu glutaminowego , Lub Dekarboksylaza glutaminianowa )] i jest katabolizowany przez inny enzym: transaminaza GABA (GABA-T).
Metabolizm GABA i glutaminianu (lub kwasu glutaminowego) są ściśle powiązane. Sam glutaminian pochodzi z glutaminy i alfa-bitoglutaranu z cyklu Krebsa (patrz ryc. 1). Glutaminian jest następnie dekarboxyle w GABA pod wpływem glutaminian -discarboksylazy (GAD) i jej kofaktoru, fosforanu pirydoksalnego, pochodzącego z witaminy B 6 .
Cząsteczki GABA są przechowywane w pęcherzykach synaptycznych przez nośniki VGAT ( pęcherzykowy transporter GABA ), utworzone przez białka transbłonowe. Przybycie potencjału czynnościowego na końcu aksonu otwiera kanały wapnia wrażliwe na napięcie. Napływ CA ++ powoduje egzocytozę, wyzwolenie GABA w szczelinie synaptycznej.
GABA wydana w szczelinie może podążać za kilkoma sposobami:
- albo zostaną schwytane przez postsynaptyczne receptory gabacinergiczne i przyczyniają się do transdukcji sygnału neuronalnego;
- być poddawane recyklingowi pośrednio przez komórkę glejową;
- albo są recyklingowe bezpośrednio przez neuron gabacinergiczny.
GABA ma zatem swój wpływ na odbiorniki lub jest regenerowane przez określonych nośników membranowych. Opisano cztery rodzaje nośników o wysokim powinowactwie [[[ 2 ] : GAT-1 w GAT-4. Nosiciele GAT-1 i -4 są bardziej wyrażane w neuronach, GAT-2 w komórkach glejowych oraz GAT-2 i 3 na poziomie nerek i wątroby.

(Ryc. 1) Metabolizm GABA.
|

(Ryc. 2) Metabolizm GABA: komórki glejowe uczestniczą w recyklingu.
|
GABA podsumowane przez neurony lub komórki glejowe jest przekształcane w sukcysynat, który wchodzi do cyklu Krebsa (zapewniające syntezę ATP). Ponieważ są to enzymy mitochondrialne (jako transferaza GABA-T), które zapewniają degradację GABA, należy je hamować, aby spowodować wzrost zawartości tkanin GABA, a zatem wzrost aktywności aktywności hamowania hamowania neurony.
Enzymy i nośniki, cele terapeutyczne [[[ modyfikator |. Modyfikator i kod ]
W niektórych zabiegach padaczkowych stosuje się właściwości hamujące ośrodkowego układu nerwowego GABA. Ta choroba neurologiczna charakteryzuje się nagłymi zrzutami nieprawidłowych impulsów nerwowych w mózgu. Rodzaj leczenia ma na celu zmniejszenie wzbudzenia neuronów poprzez zwiększenie stężenia GABA, co można wykonać poprzez hamowanie enzymów degradacji GABA-T lub transporterów recyklingu GAT:
- valproate i vigabatrine ( Sabril ), inhibitor GABA-T, który degraduje GABA;
- Tiagabina ( Gabitril ), inhibitor nośników nerwowych i gliaux GABA,
lub poprzez zwiększenie syntezy GABA, poprzez suplementację w witaminie B 6 , GAD Cofactor (enzym zapewniający syntezę GABA [[[ 2 ] ). Jest to leczenie konwulsji neo-natalnych, które jest często niewystarczające i wymaga dodania innych cząsteczek przeciwconwulsyjnych.
Receptory GABA i ich fizjologiczna rola [[[ modyfikator |. Modyfikator i kod ]
Do tej pory zidentyfikowano dwa rodzaje receptorów neuroprzekaźników:


. GABA Ionotropic Odbiorniki A składają się z 5 glikoproteicznych podjednostek, otaczających porę pozwalającą chlorom wejść, gdy GABA przywiązuje się do miejsca łącznikowego. Wejście tych anionów Cl – Hiperpolaryzacja neuronu postsynaptycznego i hamuj go [[[ 2 ] , tworząc przejście z trudniejszej fali działania:
GABA → Secucept GABA A → Wejście CL – → Hiperpolaryzacja → ↓ Potentiel D’Action
5 podjednostek każdego odbiornika pobiera z grupy 19 sklonowanych podjednostek typu α, β, γ, δ, ε, π, ρ i θ. Każda podjednostka może zawierać kilka izoform: dla α mamy α1 do α6, dla β mamy β1 do β4, dla γ z γ1 do γ3, dla ρ od ρ1 do ρ3. Większość pentamerycznych receptorów ssaków składa się z dwóch alfa, dwóch beta i gamma lub delty.
Te podjednostki odbiorników GABA A same składają się z 4 śmigieł transbłonowych z ich końcami N- i C-końcowymi zarówno pozakomórkowymi (ryc. 3 po lewej).
Biorąc pod uwagę warianty uzyskane przez alternatywne splicing, otrzymujemy znaczną liczbę wariantów, które klasyfikujemy w trzech grupach:
- Receptory GABA-A1 w GABA-A6 składają się z podjednostek α, β, γ, na przykład typ GABA-A1 składa się z α Pierwszy , B 2 , C 2 , zgodnie ze stechiometrią 2α Pierwszy 2b 2 C 2 ;
- Receptory GABA-A0 składają się z podjednostek α β δ lub α β ε. Są wrażliwe na konie, ale niewrażliwe na benzodiazepiny;
- Receptory GABA-A0R lub GABA-C składają się z powiązania ρ Pierwszy , R 2 , R 3 . Mają powinowactwo do GABA dziesięć razy wyższe niż w przypadku innych odbiorników GABA A Ale są niewrażliwe na bikukulinę i benzodiazepiny.
Zwykłe przenoszenie gabacinergiczne jest zapewnione przez receptory GABA A Znajduje się na neuronie postsynaptycznym i obejmującym podjednostkę gamma. Ale znaczenie stale aktywnego hamowania gabaminergicznego, znanego jako Tonik , nałożył się od dekady [[[ 7 ] . Najczęściej to toniczne hamowanie jest zapewniane przez receptory zawierające podjednostki delta, receptory nie obecne na poziomie synaps, ale w okolicy (zwanych odbiornikami pozadłaniowymi). Receptory zawierające podjednostki β3 i δ wykazano, że bardzo wrażliwe na GABA i etanolu. Wzmocnienie alkoholu, w pewnym sensie zależnym od dawki, przypomina subiektywne działanie zatrucia alkohol [[[ 8 ] . Badanie na temat cięć mózgu szczurów [[[ 9 ] wykazał, że alkohol zwiększa hamowanie toniczne za pośrednictwem receptorów GABA A Ekstremaptyczne (α6β3Δ) w komórkach ziarna móżdżku. Powiązane badanie behawioralne sugeruje, że alkohol wpływa na koordynację motoryczną poprzez zwiększenie tonicznego hamowania ziarnistych komórek w móżdżku.
. Odbiorniki GABA B są receptorami w połączeniu z białkiem GI/O, złożonym z siedmiu transbłonowych śmigieł. Zostały po raz pierwszy podkreślone na zakończeniach presynaptycznych [[[ 2 ] gdzie ich stymulacja powoduje zmniejszenie uwalniania neuromediatora.
Są one również obecne na poziomie somatodendrytycznym, gdzie ich stymulacja powoduje wzrost przewodnictwa potasu, związany z hiperpolaryzacją.
Odbiorniki GABA, cele terapeutyczne [[[ modyfikator |. Modyfikator i kod ]
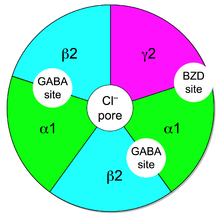
Odbiorniki GABA A są celem benzynów rodziny benzodiazepin, które wzmacniają (zwiększają) hamujący wpływ GABA na ośrodkowy układ nerwowy. I odwrotnie, niektóre karboliny beta zmniejszają hamujący działanie GABA i mają działania „pobudzające” (konwulsanci, wywołujące lęk lub pro-mnezje) w przeciwieństwie do benzodiazepin) [[[ dziesięć ] .
Miejsca łącznikowe benzodiazepin na odbiornikach GABA A są topograficznie odmienne od GABA. Te strony, powiedział allosteryczne , są rozpoznawane przez substancje czynne, które dostosują działanie GABA. Zasadniczo połączenie tych substancji modulacyjnych zwiększa częstotliwość i czas otwarcia kanału chloru, a zatem wzmacnia działanie hamujące GABA [[[ 2 ] .
Oprócz benzodiazepin znamy allosteryczne miejsca modulacyjne barbituranów, alkoholu i neurosteroidów. Wszystkie te substancje mają właściwości uspokajające, przeciwlękowe, przeciwdrgawkowe, przeciwpadaczkowe i miorelaxing.
Na odbiornikach GABA są dwa miejsca łącznikowe GABA A Znajduje się na interfejsie między podjednostkami α i β. Miejsce łącznikowej benzodiazepiny znajduje się na interfejsie między Podjednostki α i γ.
Obecność podjednostki γ jest niezbędna do rozpoznania benzodiazepin. Efekty przeciwlękowe, uspokajające i miorelusingowe tych cząsteczek są dostarczane przez receptory GABA A1, -a2, -a3 I -A5 . Z drugiej strony Zolpidem (Stilnox), rodziny blisko benzodiazepin, ma silne powinowactwo do Podjednostka α1 (I żaden dla α4, α6, δ i ε) i oddziaływania z różnymi resztami amino-acylowymi, ma wielkie powinowactwo tylko do odbiorników GABA A1 .
Cząsteczka GABA nie może przekroczyć bariery krew-mózg, dlatego rolę neuroprzekaźnika może być wykonywana tylko przez obecność GABA i uwalniana przez neurony, a nie przez wynosił żywność.
W 2016 r. Odkryto bardzo wyspecjalizowany, nazwany bakterie (jelit) (jelit) KLE1738 . Chociaż jest odległy, wydaje się, że zależy to całkowicie od biochemii mózgu w celu jego przeżycia, ponieważ spożywa tylko GABA (toksyczna cząsteczka poza pewną dawką), aby zapewnić jej metabolizm [[[ 11 ] .
Nie znamy żadnego innego jedzenia [[[ 11 ] .
Jego aktywność może częściowo wyjaśniać, w jaki sposób u ludzi (lub inne gatunki) mikrobiom jelit może wpływać na nastrój.
Niskie wskaźniki GABA były już związane z depresją i zaburzeniami nastroju; W 2011 r. Wykazaliśmy już, że różnorodność Lactobacillus (Lactobacillus rhamnosus) może znacznie zmodyfikować aktywność GABA w mózgu myszy laboratoryjnych i zmodyfikować ich odpowiedź na stres, a u myszy pozbawionych chirurgicznie nerwu błędnego (który łączy się z nerwu Vagus (który łączy się jelito bezpośrednio do mózgu) – która pozostawiła jelito do mózgu – sugerując, że w jakiś sposób odgrywa rolę w wpływie bakterii jelitowych na mózg [[[ 11 ] .
Nowe pole naukowe jest otwarte, czasem nazywane osi jelita – dla osób mówców angielskich), która teraz szuka, jeśli inne bakterie jelitowe, które konsumują lub wytwarzają GABA, w szczególności do testowania (przynajmniej u zwierząt) efektów ich Obecność w mózgu i zachowaniu, być może rozwinąć nowe leczenie przeciwko zaburzeniom nastroju (depresja, lęk, pewne formy agresywności …) [[[ 11 ] .
W 2016 r., Według badaczy INSERM, GABA może indukować regenerację komórek beta z trzustki produkującej insulinę [[[ dwunasty ] . Byłoby to przydatne szczególnie w cukrzycy typu 1, charakteryzujące się postępującą utratą tych komórek. Od tego czasu to badanie jest zaprzeczane publikacji wskazując, że wyniki nie były powtarzalne [[[ 13 ] .
Został odkryty u dwóch gatunków owadów (młoda dama i konika polna) szczyt produkcji GABA w dniu dnia w nocy, światło w kierunku zapomnienia [[[ 14 ] .
- Masa molowa obliczona po ‘ Atomowe ciężary żywiołów 2007 » , NA www.chem.qmul.ac.uk .
- Yves Landry, Jean-Pierre Gies, Docelowe farmakologię do wskazania terapeutycznego , Dunod, W 2 To jest wyd. .
- D.T. Krieger « Peptydy mózgowe: Co, gdzie i dlaczego? », Nauka W tom. 222, N O 4627, W P. 975–985 (PMID 6139875 W wznawiać ) .
- Od prążkowia (jądro caudé + putamen) do czarnej substancji Mesencephalus.
- Joaquin M. Fuster (reż.), Kora przedczołowa , Academic Press Inc, W 4 To jest wyd. , 424 P. .
- T. Sawaguchi « Odtłuszczanie cichej aktywności neuronalnej „związanej z zadaniami” w krze przedczołowej małpy przez antagonistę GABA (A) », Neuroscience Research W tom. 39, N O 1, W P. 123–131 (ISSN 0168-0102 , PMID 11164260 W Czytaj online , skonsultuałem się z )
- Ocena Farrant et Zoltan Przytulanki « Zmiany tematu hamującego: aktywacja fazowa i toniczna receptorów GABA (A) », Recenzje przyrody. Neuronauka W tom. 6, N O 3, W P. 215–229 (ISSN 1471-003X , PMID 15738957 , Doi 10.1038/NRN1625 W Czytaj online , skonsultuałem się z )
- Richard W. Olsen , Harry J. Hanchar , Pratap Meera Et Martin Wallner « Podtypy receptora GABAA: receptory „jeden kieliszek wina” », Alkohol (Fayetteville, N.Y.) W tom. 41, N O 3, W P. 201–209 (ISSN 0741-8329 , PMID 17591543 , PMCID PMC2852584 , Doi 10.1016/j.alcohol.2007.04.006 W Czytaj online , skonsultuałem się z )
- (W) H Jacob Hanchar , Paul d Dodson , Richard W. Olsen i Thomas s Otis W ‘ Upośledzenie ruchowe indukowane alkoholem spowodowane zwiększoną aktywnością receptora GABAA pozasynaptycznego GABAA » W Nature Neuroscience W tom. 8, N O 3, W P. 339–345 (ISSN 1546-1726 , Doi 10.1038/N1398 W Czytaj online , skonsultuałem się z )
- Patrice Venault i Georges Chapouth « Od behawioralnej farmakologii beta-karbolin po drgawki, lęk i pamięć », Thescientificworldjournal W tom. 7, W P. 204–223 (ISSN 1537-744X , PMID 17334612 , Doi 10.1100 / zapach.2007.48 W Czytaj online , skonsultuałem się z )
- Andy Coghlan, « Bakterie jelitowe po raz pierwszy spotykały chemikalia mózgu », New Scientist.com W ( Czytaj online ) .
- ‘ GABA, obiecująca cząsteczka leczenia cukrzycy typu 1 » , NA https://planet-vie.ens.fr .
- Amanda M. Ackermann , Nicholas G. Mech To Claus h. Kaestner « GABA i Artesunate nie indukują transdferentiacji komórek α-β-β in vivo », Metabolizm komórkowy W (ISSN 1932-7420 , PMID 30057067 , Doi 10.1016/j.cmet.2018.07.002 W Czytaj online , skonsultuałem się z )
- Raúl Vieira , Jezus M. Míguez i instrukcja Wieś « GABA moduluje zmiany w ciągu dnia w poziomach melatoniny w zwojach mózgowych damskiej Ischnura Graellsii i konika ogrodowego Oedipoda caerulescens », Litery neuronauki W tom. 376, N O 2, W P. 111–115 (ISSN 0304-3940 , PMID 15698931 , Doi 10.1016/j.neulet.2004.11.036 W Czytaj online , skonsultuałem się z )
Powiązany artykuł [[[ modyfikator |. Modyfikator i kod ]
Recent Comments