Ewolucja wici – Wikipedia
Artykuł w Wikipedii, Free L’Encyclopéi.
Zrozumienie Ewolucja płatków ma główne znaczenie dla biologów, ponieważ trzy różne eukariotyczne i prokariotyczne flagellandki są wyrafinowane i złożone struktury komórkowe pozwalające na poruszanie się komórek w ich środowisku. Ważne jest, aby określić, że flagi bakteryjne i eukariotyczne nie są homologiczne i nie mają związku.

Istnieją dwa odrębne modele dotyczące pochodzenia rzęs i flagi eukariotowej.
Model symbiotyczny [[[ modyfikator |. Modyfikator i kod ]
Model ten, zwany także endosymbiotykiem lub egzogennym, stanowi, że Gracilicutety (przodek spirochaetes i protthecobacter), w symbiozie z prymitywną archią, byłby na pochodzeniu komórki eukariotowej tych rzęs i flagelsów i flagels [[[ Pierwszy ] .
W ten sposób symbionte służyłby jako mobilny organel. Hipoteza ta jest dziełem Lynn Margulis, amerykańskiego mikrobiologa uznanego za, który stworzył i spopularyzował symbiotyczne pochodzenie mitochondriów i chloroplastów. Jednak symbiotyczne pochodzenie flagi eukariota było bardziej skrytykowane [[[ 2 ] .
Podstawowymi argumentami na korzyść teorii symbiotycznej jest to, że istnieją jednokomórkowe gatunki eukariotyczne mieszkające w jelitach zapalenia emisji, takich jak Mixotricha które mają Spirochaetes Symbiontes zapewniającą ich mobilność.
Oprócz tych przykładów symbiozy homologia między białkami wici eukariotycznej i de protthecobacter jest główną siłą tej hipotezy.
Eukariotyczne rzęsy i wici nie zawierają genomu do zidentyfikowania, w przeciwieństwie do mitochondriów i chloroplastów, ale przenoszenie tej cechy może być niezależne od jądrowego DNA.
Model endogenny [[[ modyfikator |. Modyfikator i kod ]
Ten model stanowi, że eukariotyczna rzęs i flagel pochodzą z niezbędnego cytoszkieletu eukariotycznego do tworzenia mitotycznego wrzeciona [[[ 3 ] . Białka rodziny tubulin i dynein są ważne dla tworzenia wrzeciona mitotycznego i wici eukariote. Ponadto, u niektórych eukariotycznych gatunków jednokomórkowych, podstawa eukariotowego flagellum koncentruje się i organizuje wrzeciono mitotyczne.
Potwierdzając endogenną hipotezę rzęs, badanie 3 -wymiarowej struktury tubuliny wykazało konformację podobną do białka bakteryjnego cytoszkieletu FTSz, potwierdzając w ten sposób stare filiacja początkowo w oparciu o niskie, ale znaczące homologie sekwencji [[[ 4 ] . Odkrycie to jest niezbędne, ponieważ cytoszkielet bakterii spirochaetes nie ma białka FTSZ, a zatem sugeruje, że bakterie spirochaetes nie są pochodzenia CIL.
Istnieje jednak wiele przypadków równoległej ewolucji, zwanych także zbieżną ewolucją, w których dwie cząsteczki lub dwie struktury pojawiły się niezależnie od siebie. W rezultacie struktura tubuliny może być wynikiem równoległej ewolucji [[[ 5 ] .
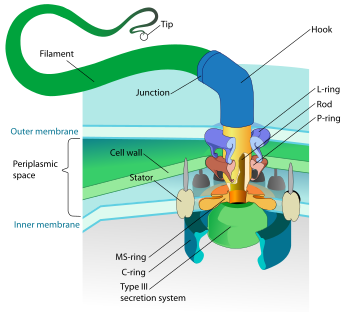
Pochodzenie bakteryjnej wici jest bardziej badane niż eukariotalna flagla. Wygląda na to, że składniki bakteryjnej flagi ewoluowały z systemu transportowego typu III, który umożliwia wydzielanie białka bakteryjnego [[[ 6 ] .
Wydzielanie białka w środowisku zewnętrznym jest złożone i obejmuje wiele kompleksów molekularnych umożliwiających celowanie i transport przez ścianę bakteryjną i błoną bakteryjną. W przypadku negatywów GRAM, które mają dwie błony, istnieje sześć systemów wydzielania o nazwie System transportu typu I do VI. System transportu typu III bierze udział w transporcie i wydzielaniu toksyn. Na przykład Bacillus Yersinia pestis , Patogen odpowiedzialny za plagę, wstrzykuje toksynę w komórkach eukariotycznych dzięki systemowi transportu typu III [[[ 7 ] . Bakteryjna flagina i system transportu III wywodzą się ze wspólnej struktury przodków, prawdopodobnie wymaganej do transportu białek. Oba systemy mają wspólne dziewięć homologicznych białek. Na przykład białko Fli1 ATPazy Fli1 jest homologiczne dla białka ATPazy T3SS (co oznacza ATPazę wydzielania typu 3). Ostatnio badania porównawcze sekwencji i struktur krystalograficznych podkreśliły silną homologię między białkiem FLIG, składnikiem silnika flagowego zlokalizowanego w dysku C oraz transporterem jonów MG2+ [[[ 8 ] .
Zwróć uwagę, aby stwierdzić, że struktura bakteryjnych wici różni się między gramem a dodatnim bakterią Grama. Rzeczywiście, podczas gdy bakterie gramowe ujemne mają dwie błony, bakterie dodatnie Gram mają tylko jedną błonę. W konsekwencji, mając inną ścianę bakteryjną i tylko jedną błonę, dodatnie bakterie Grama nie mają kompleksów stanowiących flagelię zwaną dysk P i DISC L, która w bakteriach Grama ujemnych oddziałuje odpowiednio ze ścianą bakteryjną i błoną zewnętrzną.
Analiza sekwencji białka i genetycznych ujawnia, że wicika archea jest związana z pilusem typu IV [[[ 9 ] . Flagellum archaea jest podobna, ale nie przedstawia homologii z hojami bakterii. Oprócz braku homologii sekwencji między białkami dwóch układów, flagellum archaee wzrasta przez dodanie monomeru flagelin u podstawy wici, podczas gdy dodanie błysków jest przeprowadzane w dystalnej flaginie części w bakteriach. Ponadto flagellum archaee ma mniejszą średnicę, ponieważ nie ma kanału centralnego.
Pili, dodatki pozakomórkowe, mogą cofać się i uczestniczyć w mobilności bakteryjnej za pomocą społecznego mechanizmu ślizgania się (zwanego społecznym szybowaniem lub drganiem po angielsku) poprzez interakcję z komórką gospodarza lub innymi bakteriami. Pilus typu IV jest montowany za pośrednictwem systemu transportu typu II. Na razie żadne bakterie nie używają PILI typu IV do pływania.
- Margulis, L. Chapman, M. Guerrero, R. Hall J. (2006) Ostatni eukariotyczny wspólny przodek (LECA): nabycie ruchliwości cytoszkieletowej z aerotolerancyjnych kręgetów w proterozoicznej eon Proc Natl Acad Sci U S A. 29 sierpnia; 103 (35): 13080-5
- Cavalier-Smith, T. (2006) Ewolucja komórki i historia Ziemi: zastrzeżenie i rewolucja Philos trans r soc lond b biol sci. 29 czerwca; 361 (1470): 969-1006
- Borens, M. Azimzadeh, J. (2007) Pochodzenie i ewolucja centrosomu Adv Exp Med Biol. 607: 119-29. Recenzja
- Erickson, H.P. (2007) Ewolucja cytoszkieletu Bioessays. 29 lipca; 29 (7): 668-77. Recenzja
- Edward H. Egelman (1998) Rodzina tubuliny: pokrewieństwo kluczowych białek w domenach filogenetycznych W Obecna biologia , Vol8, wydanie 8, strony R288-R290
- Pallen, J.M.; Penn, C.W .; Chaudhuri, R.R. (2005). Bakteryjna różnorodność flagowa w erze po genomowej , Trendy w Microbiology, vol.13, N4,143-149
- Shao, F. Funkcje biochemiczne efektorów Yersinia typu III CURR Opin Microbiol. 2008 lutego; 11 (1): 21-9
- Snyder, L.A.S. Loman, N.J. Fütterer, K. Pallen, M.J. Bakteryjna różnorodność i ewolucja flagelarna: Szukasz prostoty i nieufność? Trendy w Microbiology Vol.10 (1): 1-5 2009
- Faguy, DM. Jarrell, KF. Kuzio, J Kalmokoff, M.L. (1994) Analiza molekularna flagellin archaela: podobieństwo do nadransportu Pilin-Pilin-Transport w bakteriach w bakteriach , Can J Microbiol, Jan; 40 (1); 67-71 Recenzja
Recent Comments