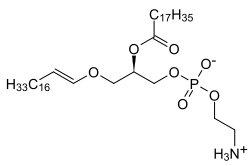
Có hai loại ether phospholipid, plasmanyl- và plasmenyl-phospholipids. Plasmanyl-phospholipids có liên kết ether ở vị trí sn-1 với nhóm alkyl. Plasmenyl-phospholipids có liên kết ether ở vị trí sn-1 với nhóm alkenyl. Loại thứ hai được gọi là plasmalogens . [1][2][3]
Ở động vật có vú, vị trí sn-1 thường có nguồn gốc từ rượu béo C16: 0, C18: 0 hoặc C18: 1 trong khi vị trí sn-2 phổ biến nhất chiếm bởi các axit béo không bão hòa đa (PUFA). Các nhóm đầu phổ biến nhất hiện diện trong plasmalogens động vật có vú là ethanolamine (plasmenylethalomines được chỉ định) hoặc choline (plasmenylcholines được chỉ định).
Chức năng [ chỉnh sửa ]
Plasmalogens được tìm thấy trong nhiều mô của con người, với sự làm giàu đặc biệt trong hệ thống thần kinh, miễn dịch và tim mạch. [1][2][3] Trong mô tim của con người, gần 30 Cấm40% choline glycerophospholipids là plasmalogens. Đáng chú ý hơn nữa là thực tế là 32% glycerophospholipids trong tim người trưởng thành và 20% trong não và tới 70% myelin vỏ bọc ethanolamine glycerophospholipids là plasmalogens. [4]
Mặc dù chức năng của plasmalogens vẫn chưa được làm sáng tỏ. người ta đã chứng minh rằng chúng có thể bảo vệ các tế bào động vật có vú chống lại tác hại của các loài oxy phản ứng. [1][2][3] Ngoài ra, chúng còn được coi là các phân tử tín hiệu và bộ điều biến của động lực màng.
Lịch sử [ chỉnh sửa ]
Plasmalogens được Feulgen và Voit mô tả lần đầu tiên vào năm 1924 dựa trên các nghiên cứu về các phần mô. [1] Họ đã xử lý các phần mô này bằng axit hoặc cloruric. một phần của phương pháp nhuộm màu hạt nhân. Điều này dẫn đến sự phá vỡ liên kết vinyl-ether plasm halogen để tạo ra aldehyd. Đổi lại, chất sau phản ứng với vết axit fuchsine-sulfurous được sử dụng trong phương pháp nhuộm hạt nhân này và làm phát sinh các hợp chất màu bên trong tế bào chất của tế bào. Plasmalogens được đặt tên dựa trên thực tế là các hợp chất có màu này có trong "plasmal" hoặc bên trong tế bào. [1]
Sinh tổng hợp [ chỉnh sửa ]

Bắt đầu sinh tổng hợp plasmalogens với sự kết hợp của các enzyme ma trận peroxisomal GNPAT (glycerone phosphate acyl transferase) và AGPS (alkyl-glycerone phosphate synthase) ở phía bên của màng peroxisomal. [5] Hai enzyme này có thể tương tác vật lý với nhau để tăng hiệu quả. Do đó, các nguyên bào sợi không có hoạt động AGPS làm giảm mức độ và hoạt động của GNPAT. [6][7]
Bước đầu tiên của quá trình sinh tổng hợp được xúc tác bởi GNPAT. Enzyme này acylates dihydroxyacetone phosphate (DHAP) ở vị trí sn-1. Tiếp theo đó là sự trao đổi nhóm acyl với nhóm alkyl bằng AGPS. [8] 1-alkyl-DHAP sau đó được khử thành 1-O-alkyl-2-hydroxy-sn-glycerophosphate (GPA) bằng acyl / alkyl-DHAP reductase nằm trong cả màng peroxisomal và retoplulum (ER). [9] Tất cả các sửa đổi khác xảy ra trong ER. Có một nhóm acyl được đặt ở vị trí sn-2 bởi một acyltransferase alkyl / acyl GPA và nhóm phosphate được loại bỏ bởi phosphatase acid phosphatase để tạo thành 1-O-alkyl-2-acyl-sn-glycerol.
Sử dụng CDP-ethanolamine một phosphotransferase tạo thành 1-O-alkyl-2-acyl-sn-GPEtn. Sau khi khử hydro ở vị trí 1- và 2 của nhóm alkyl bởi một hệ thống vận chuyển điện tử và plasmenylethanolamine desaturase liên kết vinyl ether của plasmalogens cuối cùng đã được hình thành. Plasmenylcholine được hình thành từ 1-O-alkyl-2-acyl-sn-glycerol bởi choline phosphotransferase. Vì không có plasmenylcholine desaturase choline plasmalogens chỉ có thể được hình thành sau khi thủy phân etanolamine thành 1-O- (1Z-alkenyl) -2-acyl-sn-glycerol có thể được sửa đổi bằng choline phosphotransferase và CDP choline. [ chỉnh sửa ]
Rối loạn sinh học peroxisome là rối loạn lặn tự phát thường được đặc trưng bởi sự sinh tổng hợp plasm halogen bị suy yếu. Trong những trường hợp này, enzyme peroxisomal GNPAT, cần thiết cho các bước đầu tiên của quá trình sinh tổng hợp plasm halogen, bị sai lệch với tế bào chất nơi nó không hoạt động. Ngoài ra, các đột biến gen trong các gen GNPAT hoặc AGPS có thể dẫn đến sự thiếu hụt plasm halogen, dẫn đến sự phát triển của rhizomelic chondrodysplasia puncata (RCDP) loại 2 hoặc 3. ] Trong những trường hợp như vậy, cả hai bản sao của gen GNPAT hoặc AGPS phải được biến đổi để bệnh biểu hiện. Không giống như các rối loạn sinh học peroxisome, các khía cạnh khác của sự tập hợp peroxisome ở bệnh nhân RCDP2 và RCDP3 là bình thường vì khả năng chuyển hóa các axit béo chuỗi rất dài. Các cá nhân bị thiếu hụt plasm halogen nghiêm trọng thường xuyên cho thấy sự phát triển thần kinh bất thường, dị tật xương, hô hấp bị suy yếu và đục thủy tinh thể. [13]
Sự thiếu hụt về mức độ plasm halogen góp phần vào bệnh lý trong hội chứng Zellweger. [11]
Chuột bị loại bỏ plasm halogen bắt giữ quá trình sinh tinh, phát triển đục thủy tinh thể và khiếm khuyết trong quá trình myel hóa hệ thần kinh trung ương. [14][15]
Trong quá trình viêm [ chỉnh sửa ]
Trong quá trình viêm, myeloperoxidase có nguồn gốc bạch cầu trung tính tạo ra axit hypochlorous. HOCl gây ra quá trình clo hóa oxy hóa plasmalogens ở chuỗi sn-1 bằng cách phản ứng với liên kết vinyl ether. [16] Một số nhà nghiên cứu hiện đang nghiên cứu tác động của lipid clo hóa đến bệnh lý.
Liên kết bệnh có thể xảy ra [ chỉnh sửa ]
Việc thiếu các phương pháp tốt để kiểm tra plasm halogen đã tạo ra khó khăn cho các nhà khoa học để đánh giá plasm halogen có thể liên quan đến các bệnh ở người khác ngoài phổ RCDP và Zellweger , trong đó có sự liên quan là chắc chắn. [11] Có một số bằng chứng ở người cho thấy plasmalogens thấp có liên quan đến bệnh lý của chứng loạn sản phế quản phổi, đây là một biến chứng quan trọng của việc sinh non. [11] và một nghiên cứu cho thấy mức độ plasm halogen giảm. ở những người bị COPD hút thuốc so với những người không hút thuốc. Có một số bằng chứng từ người và động vật cho thấy mức độ plasmalogens trong não bị giảm trong các rối loạn thoái hóa thần kinh bao gồm bệnh Alzheimer, bệnh Parkinson, bệnh Niemann, Pick Pick, loại C, hội chứng Down và bệnh đa xơ cứng, nhưng đến năm 2012 thì không rõ ràng nếu thâm hụt gây ra, hoặc gây ra bởi quá trình bệnh. [11]
Evolution [ chỉnh sửa ]
Ngoài động vật có vú, plasmalogens cũng được tìm thấy ở động vật không xương sống và tế bào đơn động vật nguyên sinh. Trong số các vi khuẩn, chúng đã được tìm thấy ở nhiều loài kỵ khí bao gồm Clostridia Megasphaera và Veillonella . Plasmalogens đã được chứng minh là có một lịch sử tiến hóa phức tạp dựa trên thực tế là con đường sinh tổng hợp của chúng khác nhau ở các sinh vật hiếu khí và kỵ khí. [17]
Gần đây, người ta đã chứng minh rằng các tế bào hồng cầu của con người và loài vượn lớn (tinh tinh, bonobos, khỉ đột và đười ươi) có sự khác biệt về thành phần plasm halogen của chúng. [3] Tổng mức plasm halogen của RBC được tìm thấy thấp hơn ở người so với bonobos, tinh tinh hoặc khỉ đột, nhưng cao hơn so với đười ươi. Dữ liệu biểu hiện gen từ tất cả các loài này khiến các tác giả suy đoán rằng các tế bào và mô vượn người khác và lớn khác nhau về mức độ plasm halogen. Mặc dù hậu quả của những khác biệt tiềm năng này vẫn chưa được biết rõ, sự khác biệt giữa các loài trong plasmalogens mô có thể ảnh hưởng đến chức năng cơ quan và nhiều quá trình sinh học.
Tài liệu tham khảo [ chỉnh sửa ]
- ^ a b d e Nagan, N.; Zoeller, R. A. (2001). "Plasmalogens: Sinh tổng hợp và chức năng". Tiến bộ trong nghiên cứu lipid . 40 (3): 199 Điêu229. doi: 10.1016 / S0163-7827 (01) 00003-0. PMID 11275267.
- ^ a b c Gorgas, K. Teigler, A.; Komljenovic, Đ.; Chỉ, W. W. (2006). "Chuột thiếu ether lipid: Theo dõi chức năng plasm halogen". Biochimica et Biophysica Acta (BBA) – Nghiên cứu tế bào phân tử . 1763 (12): 1511 19151526. doi: 10.1016 / j.bbamcr.2006.08.038. PMID 17027098.
- ^ a b c [199090] Moser, AB; Steinberg, S. J.; Watkins, P. A.; Moser, H. W.; Ramaswamy, K.; Siegmund, K. D..; Lee, D. R.; Ely, J. J.; Ryder, O. A.; Cây keo, J. G. (2011). "Tế bào hồng cầu của người và vượn lớn khác nhau về mức độ và thành phần plasm halogen". Lipid trong sức khỏe và bệnh tật . 10 : 101. doi: 10.1186 / 1476-511X-10-101. PMC 3129581 . PMID 21679470.
- ^ Farooqui, A. A.; Horrocks, L. A. (2001). "Plasmalogens: lipid Workhorse của màng trong tế bào thần kinh và glia bình thường và bị tổn thương". Nhà thần kinh học: một tạp chí đánh giá mang đến sinh học thần kinh, thần kinh học và tâm thần học . 7 (3): 232 Từ245. doi: 10.1177 / 107385840100700308. PMID 11499402.
- ^ P. Brites, H.R. Waterham, R.J. Người lang thang, Chức năng và sinh tổng hợp plasmalogens trong sức khỏe và bệnh tật, Biochim. Sinh lý. Acta 1636 (2004) 219 Điện231.
- ^ J. Biermann, W.W. Chỉ là, R.J. Wanders, H. Van Den Bosch, Alkyl-dihydroxyacetone phosphate synthase và dihydroxyacetone phosphate acyltransferase tạo thành một phức hợp protein trong peroxisomes, Eur. J. Sinh hóa. 261 (1999) 492 bóng499
- ^ D. Hardeman, H. van den Bosch, Địa hình sinh tổng hợp ether phospholipid, Biochim. Sinh lý. Acta 1006 (1989) 1 bóng8.
- ^ A.J. Brown, F. Snyder, Alkyldihydroxyacetone-P synthase. Sự hòa tan, tinh chế một phần, phương pháp xét nghiệm mới và bằng chứng cho một cơ chế bóng bàn, J. Biol. Hóa. 257 (1982) 8835 Từ8839
- ^ P.F. James, A.C. Lake, A.K. Hajra, L.K. Larkin, M. Robinson, F.G. Hội trưởng, R.A Zoeller, Một đột biến tế bào động vật bị thiếu hụt hoạt tính khử acyl / alkyl-dihydroxyace-tone-phosphate. Ảnh hưởng đến sinh tổng hợp liên kết ether và diacyl glycerolipids, J. Biol. Hóa. 272 (1997) 23540 Từ23546
- ^ T.C. Lee, Sinh tổng hợp và các chức năng sinh học có thể có của plasmalogens, Biochim. Sinh lý. Acta 1394 (1998) 129 Điện145
- ^ a b c ] d e Braverman, NE; Moser, AB (tháng 9 năm 2012). "Chức năng của lipid plasm halogen trong sức khỏe và bệnh tật". Biochimica et Biophysica Acta . 1822 (9): 1442 Tắt52. doi: 10.1016 / j.bbadis.2012.05.008. PMID 22627108.
- ^ Người lang thang, R.; Waterham, H. (2006). "Rối loạn peroxisomal: sự thiếu hụt enzyme peroxisomal duy nhất". Biochimica et Biophysica Acta (BBA) – Nghiên cứu tế bào phân tử . 1763 : 1707 Tiết20. doi: 10.1016 / j.bbamcr.2006.08.010. PMID 17055078.
- ^ Rhizomelic Chondrodysplasia Puncata Loại 1. Tác giả Braverman NE, Moser AB, Steinberg SJ. Biên tập viên trong: Pagon RA, Adam MP, Bird TD, Dolan CR, Fong CT, Smith RJH, Stephens K, biên tập viên. Nguồn GeneReview [Internet]. Seattle (WA): Đại học Washington, Seattle; 1993-2014. 2001 16 tháng 11
- ^ Gorgas K, Teigler A, Komljenovic D, Just WW. Chuột thiếu ether lipid: theo dõi các chức năng plasm halogen.Biochim Biophys Acta. 2006 Dec; 1763 (12): 1511-26
- ^ Rodemer C, Thai TP, Brugger B, Kaercher T, Werner H, Nave KA, Wieland F, Gorgas K, Just WW. Bất hoạt sinh tổng hợp lipid ether gây vô sinh nam, khiếm khuyết trong phát triển mắt và giảm sản thần kinh thị giác ở chuột.Hum Mol Genet. 2003 1 tháng 8; 12 (15): 1881-95.
- ^ Albert, Carolyn J.; Crowley, Jan R.; Hsu, Fong-Fu; Thukkani, Arun K.; Ford, David A. (2001-06-29). "Các loài clo phản ứng được sản xuất bởi Myeloperoxidase Nhắm vào liên kết Vinyl Ether của Plasmalogens NHẬN DẠNG 2-CHLOROHEXADECANAL". Tạp chí Hóa học sinh học . 276 (26): 23733 Cách23741. doi: 10.1074 / jbc.M101447200. ISSN 0021-9258. PMID 11301330.
- ^ Goldfine, H. (2010). "Sự xuất hiện, biến mất và xuất hiện trở lại của plasmalogens trong quá trình tiến hóa". Tiến bộ trong nghiên cứu lipid . 49 (4): 493 Tiết498. doi: 10.1016 / j.plipres.2010.07.003. PMID 20637230.
Liên kết ngoài [ chỉnh sửa ]