TGF Beta-Signalweg – Wikipedia
Signalweg mit transformierenden Beta-Proteinen des Wachstumsfaktors
Das Signalweg des transformierenden Wachstumsfaktors Beta (TGFB) ist an vielen zellulären Prozessen sowohl im adulten Organismus als auch im sich entwickelnden Embryo beteiligt, einschließlich Zellwachstum, Zelldifferenzierung, Apoptose, zellulärer Homöostase und anderen zellulären Funktionen. Trotz der Vielzahl von zellulären Prozessen, die der TGFβ-Signalweg reguliert, ist der Prozess relativ einfach. Liganden der TGFβ-Superfamilie binden an einen Typ-II-Rezeptor, der einen Typ-I-Rezeptor rekrutiert und phosphoryliert. Der Typ I-Rezeptor phosphoryliert dann rezeptorregulierte SMADs (R-SMADs), die nun das coSMAD SMAD4 binden können. R-SMAD / coSMAD-Komplexe reichern sich im Kern an, wo sie als Transkriptionsfaktoren wirken und an der Regulation der Zielgenexpression beteiligt sind.
Mechanismus[edit]
Ligandenbindung[edit]
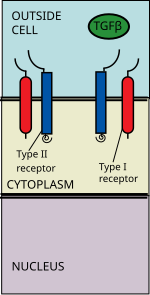
Die TGF-Beta-Superfamilie der Liganden umfasst: Knochenmorphogenetische Proteine (BMPs), Wachstums- und Differenzierungsfaktoren (GDFs), Anti-Müller-Hormon (AMH), Activin, Nodal und TGFβ.[1] Die Signalübertragung beginnt mit der Bindung eines Liganden der TGF-Beta-Superfamilie an einen TGF-Beta-Typ-II-Rezeptor. Der Typ II-Rezeptor ist eine Serin / Threonin-Rezeptorkinase, die die Phosphorylierung des Typ I-Rezeptors katalysiert. Jede Ligandenklasse bindet an einen bestimmten Typ-II-Rezeptor.[2] Bei Säugetieren sind sieben Typ I-Rezeptoren und fünf Typ II-Rezeptoren bekannt.[3]
Es gibt drei Aktivine: Activin A, Activin B und Activin AB. Activine sind an der Embryogenese und Osteogenese beteiligt. Sie regulieren auch viele Hormone, einschließlich Hypophysen-, Gonaden- und Hypothalamushormone sowie Insulin. Sie sind auch Überlebensfaktoren für Nervenzellen.
Die BMPs binden an den knochenmorphogenetischen Proteinrezeptor Typ 2 (BMPR2). Sie sind an einer Vielzahl von Zellfunktionen beteiligt, einschließlich Osteogenese, Zelldifferenzierung, Spezifikation der Vorder- / Hinterachse, Wachstum und Homöostase.
Die TGF-Beta-Familie umfasst: TGFβ1, TGFβ2, TGFβ3. Wie die BMPs sind TGF-Betas an der Embryogenese und Zelldifferenzierung beteiligt, aber sie sind auch an der Apoptose sowie an anderen Funktionen beteiligt. Sie binden an den TGF-beta-Rezeptor Typ 2 (TGFBR2).
Nodal bindet an den Aktivin A-Rezeptor Typ IIB ACVR2B. Es kann dann entweder einen Rezeptorkomplex mit Activin A-Rezeptor Typ IB (ACVR1B) oder mit Activin A-Rezeptor Typ IC (ACVR1C) bilden.[3]
Wenn die Rezeptor-Ligand-Bindung über lokale Wirkung erfolgt, wird dies als parakrine Signalübertragung klassifiziert.
Rezeptorrekrutierung und Phosphorylierung[edit]

Der TGF-Beta-Ligand bindet an ein Typ-II-Rezeptor-Dimer, das ein Typ-I-Rezeptor-Dimer rekrutiert und mit dem Liganden einen heterotetrameren Komplex bildet.[4] Diese Rezeptoren sind Serin / Threonin-Kinase-Rezeptoren. Sie haben eine cysteinreiche extrazelluläre Domäne, eine Transmembrandomäne und eine cytoplasmatische Serin / Threonin-reiche Domäne. Die GS-Domäne des Typ I-Rezeptors besteht aus einer Reihe von etwa dreißig Serin-Glycin-Wiederholungen.[5] Die Bindung eines Liganden der TGF-Beta-Familie bewirkt die Rotation der Rezeptoren, so dass ihre cytoplasmatischen Kinasedomänen in einer katalytisch günstigen Orientierung angeordnet sind. Der Typ II-Rezeptor phosphoryliert Serinreste des Typ I-Rezeptors, der das Protein aktiviert.
SMAD-Phosphorylierung[edit]

Es gibt fünf rezeptorregulierte SMADs: SMAD1, SMAD2, SMAD3, SMAD5 und SMAD9 (manchmal auch als SMAD8 bezeichnet). Es gibt im Wesentlichen zwei intrazelluläre Wege, an denen diese R-SMADs beteiligt sind. TGF-Betas, Activine, Nodals und einige GDFs werden durch SMAD2 und SMAD3 vermittelt, während BMPs, AMH und einige GDFs durch SMAD1, SMAD5 und SMAD9 vermittelt werden. Die Bindung des R-SMAD an den Typ I-Rezeptor wird durch eine Protein enthaltende Zink-Doppelfinger-FYVE-Domäne vermittelt. Zwei solche Proteine, die den TGF-Beta-Weg vermitteln, umfassen SARA (der SMAD-Anker für die Rezeptoraktivierung) und HGS (Hepatozytenwachstumsfaktor-reguliertes Tyrosinkinasesubstrat).
SARA ist in einem frühen Endosom vorhanden, das durch Clathrin-vermittelte Endozytose den Rezeptorkomplex internalisiert.[6]SARA rekrutiert einen R-SMAD. SARA ermöglicht die Bindung des R-SMAD an die L45-Region des Typ I-Rezeptors.[7] SARA richtet das R-SMAD so aus, dass der Serinrest an seinem C-Terminus der katalytischen Region des Typ I-Rezeptors zugewandt ist. Der Typ I-Rezeptor phosphoryliert den Serinrest des R-SMAD. Die Phosphorylierung induziert eine Konformationsänderung in der MH2-Domäne des R-SMAD und dessen anschließende Dissoziation vom Rezeptorkomplex und SARA.[8]
CoSMAD-Bindung[edit]

Das phosphorylierte RSMAD hat eine hohe Affinität zu einem coSMAD (z. B. SMAD4) und bildet mit einem einen Komplex. Die Phosphatgruppe fungiert nicht als Andockstelle für coSMAD, sondern die Phosphorylierung eröffnet eine Aminosäurestrecke, die eine Wechselwirkung ermöglicht.
Transkription[edit]

Der phosphorylierte RSMAD / coSMAD-Komplex tritt in den Kern ein, wo er Transkriptionspromotoren / Cofaktoren bindet und die Transkription von DNA bewirkt.
Knochenmorphogenetische Proteine verursachen die Transkription von mRNAs, die an der Osteogenese, Neurogenese und ventralen Mesodermspezifikation beteiligt sind.
TGF-Betas verursachen die Transkription von mRNAs, die an Apoptose, extrazellulärer Matrixneogenese und Immunsuppression beteiligt sind. Es ist auch am G1-Arrest im Zellzyklus beteiligt.
Activin bewirkt die Transkription von mRNAs, die am Gonadenwachstum, der Embryodifferenzierung und der Plazentabildung beteiligt sind.
Nodal bewirkt die Transkription von mRNAs, die an der Spezifikation der linken und rechten Achse, der Mesoderm- und Endoderm-Induktion beteiligt sind.
Wegregulierung[edit]
Der TGF-Beta-Signalweg ist an einer Vielzahl von zellulären Prozessen beteiligt und wird anschließend sehr stark reguliert. Es gibt eine Vielzahl von Mechanismen, bei denen der Signalweg entweder positiv oder negativ moduliert wird: Es gibt Agonisten für Liganden und R-SMADs; es gibt Täuschungsrezeptoren; und R-SMADs und Rezeptoren sind ubiquitiniert.
Ligandenagonisten / -antagonisten[edit]
Sowohl Chordin als auch Noggin sind Antagonisten von BMPs. Sie binden BMPs, die die Bindung des Liganden an den Rezeptor verhindern.[9] Es wurde gezeigt, dass Chordin und Noggin das Mesoderm dorsalisieren. Sie befinden sich beide in der Rückenlippe von Xenopus und ansonsten epidermisspezifisches Gewebe in Nervengewebe umwandeln (siehe Neurulation). Noggin spielt eine Schlüsselrolle bei der Knorpel- und Knochenstrukturierung. Mäuse Noggin – / – haben überschüssigen Knorpel und keine Gelenkbildung.[9]
Mitglieder der DAN-Proteinfamilie antagonisieren auch Mitglieder der TGF-Beta-Familie. Dazu gehören Cerberus, DAN und Gremlin. Diese Proteine enthalten neun konservierte Cysteine, die Disulfidbrücken bilden können. Es wird angenommen, dass DAN GDF5, GDF6 und GDF7 antagonisiert.
Follistatin hemmt Activin, das es bindet. Es wirkt sich direkt auf die Sekretion des follikelstimulierenden Hormons (FSH) aus. Follistatin ist auch an Prostatakrebs beteiligt, bei dem Mutationen in seinem Gen verhindern können, dass es auf Aktivin mit antiproliferativen Eigenschaften einwirkt.[9]
Lefty ist ein Regulator von TGFβ und an der Achsenstrukturierung während der Embryogenese beteiligt. Es ist auch ein Mitglied der TGF-Superfamilie der Proteine. Es wird asymmetrisch auf der linken Seite von Mausembryonen exprimiert und spielt anschließend eine Rolle bei der Links-Rechts-Spezifikation. Lefty verhindert die Phosphorylierung von R-SMADs. Dies geschieht durch einen konstitutiv aktiven TGFβ-Typ-I-Rezeptor und durch einen Prozess stromabwärts seiner Aktivierung.[10]
Es wurden auch Antagonisten auf Arzneimittelbasis identifiziert, wie SB431542,[11] das selektiv ALK4, ALK5 und ALK7 hemmt.
Rezeptorregulierung[edit]
Der transformierende Wachstumsfaktor-Rezeptor 3 (TGFBR3) ist der bisher am häufigsten vorkommende der TGF-β-Rezeptoren.[12] Es ist keine Signalisierungsdomäne bekannt.[13] Es kann jedoch dazu dienen, die Bindung von TGF-beta-Liganden an TGF-beta-Typ-II-Rezeptoren zu verstärken, indem TGFβ gebunden und an TGFBR2 präsentiert wird. Eines der nachgeschalteten Ziele der TGF-β-Signalübertragung, GIPCbindet an seine PDZ-Domäne, wodurch sein proteosomaler Abbau verhindert wird, der anschließend die TGFβ-Aktivität erhöht. Es kann auch als Inhibin-Coreceptor für ActivinRII dienen.[9]
BMP und Activin Membrane Bound Inhibitor (BAMBI) haben eine ähnliche extrazelluläre Domäne wie Typ I-Rezeptoren. Es fehlt eine intrazelluläre Serin / Threonin-Proteinkinasedomäne und ist daher ein Pseudorezeptor. Es bindet an den Typ I-Rezeptor und verhindert so dessen Aktivierung. Es dient als negativer Regulator der TGF-Beta-Signalübertragung und kann die tgf-Beta-Expression während der Embryogenese einschränken. Für seine Expression ist eine BMP-Signalisierung erforderlich
FKBP12 bindet die GS-Region des Typ I-Rezeptors und verhindert die Phosphorylierung des Rezeptors durch die Typ II-Rezeptoren. Es wird angenommen, dass FKBP12 und seine Homologen dazu beitragen, die Typ I-Rezeptoraktivierung in Abwesenheit eines Liganden zu verhindern, da die Ligandenbindung seine Dissoziation verursacht.
R-SMAD-Regulierung[edit]
Rolle inhibitorischer SMADs[edit]
Es gibt zwei weitere SMADs, die die SMAD-Familie vervollständigen, die inhibitorischen SMADs (I-SMADS), SMAD6 und SMAD7. Sie spielen eine Schlüsselrolle bei der Regulierung der TGF-Beta-Signalübertragung und sind an negativen Rückkopplungen beteiligt. Wie andere SMADs haben sie eine MH1- und eine MH2-Domäne. SMAD7 konkurriert mit anderen R-SMADs mit dem Typ I-Rezeptor und verhindert deren Phosphorylierung.[9][14] Es befindet sich im Kern und wandert nach Aktivierung des TGF-Beta-Rezeptors in das Zytoplasma, wo es den Typ I-Rezeptor bindet. SMAD6 bindet SMAD4 und verhindert so die Bindung anderer R-SMADs an das coSMAD. Die I-SMAD-Spiegel steigen mit der TGF-Beta-Signalgebung an, was darauf hindeutet, dass sie nachgeschaltete Ziele der TGF-Beta-Signalgebung sind.
R-SMAD-Ubiquitinierung[edit]
Die E3-Ubiquitin-Protein-Ligasen SMURF1 und SMURF2 regulieren die SMAD-Spiegel. Sie akzeptieren Ubiquitin von einem E2-konjugierenden Enzym, wo sie Ubiquitin auf die RSMADs übertragen, was ihre Ubiquitinierung und den anschließenden proteosomalen Abbau verursacht. SMURF1 bindet an SMAD1 und SMAD5, während SMURF2 SMAD1, SMAD2, SMAD3, SMAD6 und SMAD7 bindet. Es[clarification needed] verstärkt die inhibitorische Wirkung von SMAD7, während die Transkriptionsaktivitäten von SMAD2 reduziert werden.
Übersichtstabelle[edit]
TGF-β-Liganden von H. sapiens grau hervorgehoben, von D. melanogaster Liganden in rosa, von C.elegans in Gelb.
| Ligand der TGF-β-Superfamilie | Ligandeninhibitoren | Typ II Rezeptor | Typ I Rezeptor | R-SMADs | coSMAD | I-SMADs |
|---|---|---|---|---|---|---|
| Activin A. | Follistatin | ACVR2A | ACVR1B (ALK4) | SMAD2, SMAD3 | SMAD4 | SMAD7 |
| GDF1 | ACVR2A | ACVR1B (ALK4) | SMAD2, SMAD3 | SMAD4 | SMAD7 | |
| GDF11 | ACVR2B | ACVR1B (ALK4), TGFβRI (ALK5) | SMAD2, SMAD3 | SMAD4 | SMAD7 | |
| BMP2-8 | Noggin, Chordin, DAN | BMPR2 | BMPR1A (ALK3), BMPR1B (ALK6) | SMAD1 SMAD5, SMAD8 | SMAD4 | SMAD6, SMAD7 |
| Nodal | Lefty | ACVR2B | ACVR1B (ALK4), ACVR1C (ALK7) | SMAD2, SMAD3 | SMAD4 | SMAD7 |
| TGFβs | LTBP1, THBS1, Decorin | TGFβRII | TGFβRI (ALK5) | SMAD2, SMAD3 | SMAD4 | SMAD7 |
| Dpp | Punt | Tkv | Wütend | Medea | ||
| Schraube | Punt | Saxophon | Wütend | Medea | ||
| Myoglianin | Witz | Pavian | dSmad2 | Medea | ||
| dActivin | Esprit, Punt | Pavian | dSmad2 | Medea | ||
| Gbb | Esprit, Punt | Tkv, Sax | Wütend | Medea | ||
| Daf-7 | Daf-4 | Daf-1 | Daf-8, Daf-14 | Daf-3 | ||
| Dbl-1 | Daf-4 | Sma-6 | Sma-2, Sma-3, Sma-4 | Sma-4 |
Externe Links[edit]
Verweise[edit]
- ^ “Prosite-Dokumentation PDOC00223”. Archiviert von das Original am 25.05.2011. Abgerufen 2006-07-01.
- ^ Alberts B., Johnson A., Lewis J., Raff M., Roberts K., Walter P. (2002). Molekularbiologie der Zelle. New York, NY: Garland Science. ISBN 978-0-8153-3218-3.
- ^ ein b Munir S., Xu G., Wu Y, Yang B., Lala PK, Peng C. (Juli 2004). “Nodal und ALK7 hemmen die Proliferation und induzieren Apoptose in menschlichen Trophoblastenzellen.”. Das Journal of Biological Chemistry. 279 (30): 31277–86. doi:10.1074 / jbc.M400641200. PMID 15150278.
- ^ Wrana JL, Attisano L., Cárcamo J., Zentella A., Doody J., Laiho M. et al. (Dezember 1992). “TGF-Beta-Signale durch einen heteromeren Proteinkinase-Rezeptorkomplex”. Zelle. 71 (6): 1003–14. doi:10.1016 / 0092-8674 (92) 90395-S. PMID 1333888.
- ^ “Pfam Eintrag TGF_beta_GS”. Abgerufen 2006-07-01.
- ^ Runyan CE, Schnaper HW, Poncelet AC (März 2005). “Die Rolle der Internalisierung bei der Transformation der Beta1-induzierten Smad2-Assoziation des Wachstumsfaktors mit dem Smad-Anker für die Rezeptoraktivierung (SARA) und die Smad2-abhängige Signalübertragung in menschlichen Mesangialzellen”. Das Journal of Biological Chemistry. 280 (9): 8300–8. doi:10.1074 / jbc.M407939200. PMID 15613484.
- ^ Moustakas A (September 2002). “Smad Signalisierungsnetzwerk”. Journal of Cell Science. 115 (Pt 17): 3355–6. PMID 12154066.
- ^ Souchelnytskyi S., Rönnstrand L., Heldin CH, Ten Dijke P. (2001). “Phosphorylierung von Smad-Signalproteinen durch Rezeptor-Serin / Threonin-Kinasen”. Proteinkinase-Protokolle. Methoden der Molekularbiologie. 124. S. 107–20. doi:10.1385 / 1-59259-059-4: 107. ISBN 1-59259-059-4. PMID 11100470.
- ^ ein b c d e Massagué J, Chen YG (März 2000). “Steuerung der TGF-beta-Signalübertragung”. Gene & Entwicklung. 14 (6): 627–44. PMID 10733523.
- ^ Ulloa L, Tabibzadeh S. (Juni 2001). “Lefty hemmt die Rezeptor-regulierte Smad-Phosphorylierung, die durch den aktivierten transformierenden Wachstumsfaktor-Beta-Rezeptor induziert wird.”. Das Journal of Biological Chemistry. 276 (24): 21397–404. doi:10.1074 / jbc.M010783200. PMID 11278746.
- ^ Laping NJ, Grygielko E., Mathur A., Butter S., Bomberger J., Tweed C. et al. (Juli 2002). Hemmung der durch transformierenden Wachstumsfaktor (TGF) -beta1 induzierten extrazellulären Matrix mit einem neuen Inhibitor der TGF-beta-Typ-I-Rezeptorkinaseaktivität: SB-431542. Molekulare Pharmakologie. 62 (1): 58–64. doi:10.1124 / mol.62.1.58. PMID 12065755.
- ^ Blobe GC, Liu X, Fang SJ, How T, Lodish HF (Oktober 2001). “Ein neuartiger Mechanismus zur Regulierung der Signalübertragung des transformierenden Wachstumsfaktors Beta (TGF-beta). Funktionelle Modulation der TGF-beta-Rezeptorexpression vom Typ III durch Wechselwirkung mit dem PDZ-Domänenprotein GIPC”. Das Journal of Biological Chemistry. 276 (43): 39608–17. doi:10.1074 / jbc.M106831200. PMID 11546783.
- ^ Online-Mendelsche Vererbung beim Menschen (OMIM): TRANSFORMING WACHSTUMSFAKTOR-BETA-EMPFÄNGER, TYP III; TGFBR3 – 600742
- ^ Itoh F., Asao H., Sugamura K., Heldin CH, Ten Dijke P., Itoh S. (August 2001). “Förderung der Signalübertragung von morphogenetischem Knochenprotein durch negative Regulation inhibitorischer Smads”. Das EMBO Journal. 20 (15): 4132–42. doi:10.1093 / emboj / 20.15.4132. PMC 149146. PMID 11483516.
Recent Comments