Tsintaosaurus – Wikipedia
Hadrosaurid Ornithopod Dinosaurier aus der späten Kreidezeit
Tsintaosaurus (; bedeutet “Qingdao-Eidechse” nach der alten Transliteration “Tsingtao”) [2] ist eine Gattung von Hadrosauriden aus China. Es war ungefähr 8,3 Meter lang und wog 2,5 Tonnen.[3] Die Typusart ist Tsintaosaurus spinorhinus, erstmals 1958 vom chinesischen Paläontologen CC Young beschrieben.
Ein Hadrosaurier, Tsintaosaurus hatte eine charakteristische Schnauze aus Entenschnabel und eine Batterie kräftiger Zähne, mit denen die Vegetation gekaut wurde. Es ging normalerweise auf allen vieren, konnte sich aber auf den Hinterbeinen aufrichten, um nach Raubtieren zu suchen und zu fliehen, wenn es eines entdeckte. Wie andere Hadrosaurier, Tsintaosaurus lebte wahrscheinlich in Herden.
Entdeckung und Benennung[edit]

1950 wurden in Hsikou bei Chingkangkou in Laiyang, Shandong im Osten Chinas verschiedene Überreste großer Hadrosauriden entdeckt. 1958 wurden diese vom chinesischen Paläontologen Yang Zhongjian (“CC Young”) als Typusart beschrieben Tsintaosaurus spinorhinus. Der Gattungsname leitet sich von der Stadt Qingdao ab, die früher oft als “Tsintao” übersetzt wurde. Der spezifische Name bedeutet aus dem Lateinischen “mit einem Nasenrücken” Spinaund Griechisch ῥίς, Rhis, “Nase”, in Bezug auf das markante Wappen auf der Schnauze.[1]
Der Holotyp IVPP AS V725 wurde in einer Schicht der Jingangkou-Formation entdeckt, die Teil der Wangshi-Gruppe aus dem Campanian ist. Es besteht aus einem Teilskelett mit Schädel. Der Paratyp ist das Exemplar IVPP V818, ein Schädeldach. Im gleichen Gebiet wurden einige zusätzliche Teilskelette und eine große Anzahl disartikulierter Skelettelemente gefunden. Einige davon wurden von Yang erwähnt Tsintaosaurusandere wurden als benannt Tanius chingkankouensis Yang 1958; auch ein Tanius laiyangensis Zhen 1976 existiert. Die beiden letztgenannten Arten gelten heute entweder als Junior-Synonyme oder Nomina Dubia. Spätere Forscher würden einen größeren Teil des Materials auf verweisen Tsintaosaurus.
Beschreibung[edit]
Kamm[edit]

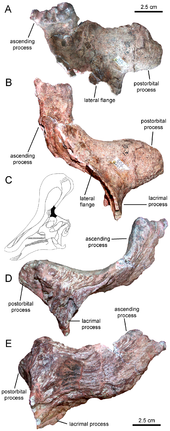
Tsintaosaurus wurde ursprünglich mit einem einhornartigen Wappen auf dem Schädel rekonstruiert. Das erhaltene Wappen besteht aus einem etwa vierzig Zentimeter langen Prozess, der fast senkrecht aus der Oberseite der hinteren Schnauze herausragt. Die Struktur ist hohl und scheint ein gegabeltes oberes Ende zu haben. Vergleichbare Strukturen mit verwandten Arten sind unbekannt: Sie besitzen mehr lappenartige Kämme. Im Jahr 1990 bezweifelten David Weishampel und Jack Horner das Vorhandensein des Kamms, was darauf hindeutete, dass es sich tatsächlich um einen gebrochenen Nasenknochen von der Spitze der Schnauze handelte, der durch ein Quetschen des Fossils nach oben verzerrt wurde. Ihre Studie legte ferner nahe, dass ohne das charakteristische Wappen, um es zu unterscheiden, Tsintaosaurus war eigentlich ein Synonym für den ähnlichen, aber kammlosen Hadrosaurier Tanius. 1993 gelangte Eric Buffetaut ea nach einer erneuten Untersuchung der Knochen selbst zu dem Schluss, dass das Wappen weder verzerrt noch ein Artefakt der Restaurierung war. Außerdem war seitdem ein zweites Exemplar mit einem aufrechten Kammteil entdeckt worden, was darauf hinweist, dass das Kamm tatsächlich echt war und Tsintaosaurus ist wahrscheinlich eine bestimmte Gattung.[4]

Eine neue Rekonstruktion im Jahr 2013 von Albert Prieto-Márquez und Jonathan Wagner, basierend auf der Identifizierung des Exemplars IVPP V829, einer Praemaxilla, als Tsintaosaurus Element kam zu dem Schluss, dass der einhornartige Knochen nur der hintere Teil eines größeren Schädelkamms war, der von der Spitze der Schnauze aus begann. Die Vorderseite des Kamms wäre durch aufsteigende Prozesse der Praemaxillen gebildet worden. Diese hatten rhomboide Kontaktfacetten mit den erweiterten oberen Teilen der Kammfortsätze der Nasenknochen erweitert und bildeten die Rückseite des Kamms. Die hintere Basis des Kamms war von Auswüchsen der Präfrontalen bedeckt. Die verschmolzenen Nasenknochen hätten eine hohle röhrenförmige Struktur gebildet. Die Höhe des Kamms hätte die des hinteren Schädels überschritten, gemessen entlang der Quadrate. Obwohl weitgehend vertikal, ist der Kamm leicht nach hinten gerichtet; Die Vorwärtsneigung des Holotypkamms wäre das Ergebnis einer Verzerrung des Fossils.[5]

Die neue Rekonstruktion von Prieto-Márquez und Wagner führte auch zu einer neuen Hypothese über die inneren Luftwege des Kamms. Yang hatte angenommen, dass die röhrenförmige Vertiefung im erhaltenen Teil des Holotyps als Hauptluftansaugung gedient hätte. Dies wurde von Prieto-Márquez und Wagner abgelehnt, die darauf hinwiesen, dass die Röhre an ihrem unteren Ende geschlossen war und dass sich bei Lambeosaurinen die Luftwege im Allgemeinen in einer weiter vorne liegenden Position befinden, wobei die knöchernen Nasenlöcher vollständig von den Praemaxillen umschlossen sind. Das haben sie angenommen Tsintaosaurus hätte eine Standard-Lambeosaurin-Anordnung in der Schnauze gehabt, die Luft, beim Einatmen, in den Schädel durch das Paar eingedrungen Pseudonares, die “falschen Nasenlöcher” der Praemaxillen hinter dem oberen Schnabel. Von dort wäre die Luft durch paarweise Passagen unterhalb der Medianfortsätze der Praemaxillen zur Spitze des Kamms transportiert worden und anschließend in eine gemeinsame Median-Kammer innerhalb des Lappens eingetreten. Der hintere Teil der Kammer wurde von den Nasenknochen gebildet und war wahrscheinlich homolog zur Nasenhöhle. Die Kammer wurde durch gekrümmte Mittelfortsätze der Praemaxillen in zwei kleinere Hohlräume unterteilt, einen vorne und einen hinten, wobei Haken um einen Durchgang zwischen den Hohlräumen gebildet wurden. Von der hinteren Höhle wurde die Luft nach unten in Richtung der inneren Schädelhöhle transportiert. Obwohl normalerweise angenommen wird, dass eine einzige Passage für diesen Zweck diente, sahen Prieto-Márquez und Wagner Hinweise in Form der Nase, dass es nach unten gepaarte Passagen innerhalb der lateralen Prozesse der Praemaxillen gab. Daraus folgerten sie, dass der gesamte Luftstrom wahrscheinlich getrennt war, wobei die Kammer des gemeinsamen Mediums wahrscheinlich durch ein knorpeliges Septum in einen linken und einen rechten Abschnitt unterteilt war.[5]
Die Schlussfolgerung, dass die röhrenförmige Struktur in den hinteren Nasenknochen kein Luftdurchgang war, zwang Prieto-Márquez und Wagner, eine alternative Erklärung für ihre Funktion zu finden. Sie schlugen vor, dass es dazu gedient hätte, das Gewicht des Kamms zu verringern, ein solches Rohr, das relative Stärke mit einer geringen Knochenmasse kombiniert. Tsintaosaurus hätte sich darin von mehr abgeleiteten Lambeosaurinen unterschieden, die eine vordere Verlängerung des Stirnknochens in Form eines Knochenblatts haben, das den Kamm stützt.[5]
Andere charakteristische Merkmale[edit]

Neben dem Wappen identifizierten Prieto-Márquez und Wagner mehrere andere charakteristische Merkmale (Autapomorphien) von Tsintaosaurus. Der Rand des oberen Schnabels ist abgerundet und dick und breiter als die Querbreite der vorderen Vertiefung um die Nasenlöcher. Soweit sich diese Vertiefung auf den Praemaxillen befindet, ist sie auf jeder Seite in Längsrichtung durch zwei Grate geteilt, die schräg nach unten und seitlich verlaufen. Intern bilden die verschmolzenen Nasenknochen einen knöchernen Block vor der Gehirnschale. Die Rückseite des Nasenknochens wird durch vordere Verlängerungen des Frontalknochens abgeschnitten, von denen die oberste relativ zum Schädeldach angehoben ist. Die aufsteigenden Äste der Praemaxillen weisen interne Prozesse auf, die nach hinten, unten und leicht nach innen zeigen und eine gemeinsame Kammer in der Mittellinie teilen. Das Präfrontale besitzt einen Flansch, der sich vom unteren Teil des Tränenknochens zum unteren Teil des aufsteigenden Prozesses des Präfrontalen erstreckt und mit einem Prozess an der Seite der Praemaxilla verbunden ist, um eine Erhebung an der Seite der Kammbasis zu bilden. Die Seite und die Unterseite des Präfrontalen zeigen tiefe vertikale Rillen. Das supratemporale Fenster ist quer breiter als lang.[5]
Einstufung[edit]
Tsintaosaurus kann eine Klade in Lambeosaurinae mit den europäischen Gattungen bilden Pararhabdodon und Koutalisaurus (wahrscheinliches Synonym von Pararhabdodon).[6]
Die Position von Tsintaosaurus im Evolutionsbaum nach einer Studie von Prieto-Márquez aus dem Jahr 2013 wird ea durch dieses Cladogramm angezeigt:[7]
Paläoökologie[edit]
Eine Untersuchung von Dinosaurier-Eiern in aufeinanderfolgenden Schichten der Wangshi-Serie der Provinz Shandong, von denen die Jingangkou-Formation die jüngste Schicht ist, zeigt, dass die Region eine hohe Dinosauriervielfalt aufweist und das Klima gegenüber der vorhergehenden Jiangjunding-Formation trockener geworden ist.[8]
Siehe auch[edit]
Verweise[edit]
- ^ ein b c Young, C.-C. (1958). “Die Dinosaurierreste von Laiyang, Shantung”. Palaeontologia Sinica, Neue Serie C.. Ganze Zahl. 42 (16): 1–138.
- ^ Creisler, B. (2002). “Dinosauria Translation and Aussprache Guide T”. DOL Dinosaurier Omnipedia. Archiviert von das Original am 31. Dezember 2005. Abgerufen 24. Februar 2010.
- ^ Gregory S. Paul (2010). Der Prince Field Guide für Dinosaurier. Vereinigte Staaten: Princeton University Press. pp. 308. ISBN 978-0-691-13720-9.
- ^ Buffetaut, E.; Tong, H. (1993). “”Tsintaosaurus spinorhinus Jung und Tanius sinensis Wiman: eine vorläufige vergleichende Studie von zwei Hadrosauriern (Dinosauria) aus der Oberkreide Chinas “. 2. 317. CR Academy of Science Paris: 1255–1261.
- ^ ein b c d Prieto-Márquez, A.; Wagner, JR (2013). “Der ‘Einhorn’-Dinosaurier, der es nicht war: Eine neue Rekonstruktion des Wappens von Tsintaosaurus und die frühe Entwicklung des Lambeosaurine Crest and Rostrum “. PLUS EINS. 8 (11): e82268. Bibcode:2013PLoSO … 882268P. doi:10.1371 / journal.pone.0082268. PMC 3838384. PMID 24278478.
- ^ Prieto-Márquez, A.; Wagner, JR (2009). “”Pararhabdodon isonensis und Tsintaosaurus spinorhinus: eine neue Gruppe von Lambeosaurin-Hadrosauriden aus Eurasien “. Kreideforschung. Online-Preprint (5): 1238–1246. doi:10.1016 / j.cretres.2009.06.005. hdl:2152/41080.
- ^ Prieto-Marquez, A.; Vecchia, FMD; Gaete, R.; Galobart, A. (2013). “Vielfalt, Beziehungen und Biogeographie der Lambeosaurin-Dinosaurier aus dem europäischen Archipel mit Beschreibung des neuen Aralosaurins Canardia garonnensis“”. PLUS EINS. 8 (7): e69835. Bibcode:2013PLoSO … 869835P. doi:10.1371 / journal.pone.0069835. PMC 3724916. PMID 23922815.
- ^ Zhao, ZiKui; Zhang, ShuKang; Wang, Qiang; Wang, XiaoLin (2013). “Dinosauriervielfalt während des Übergangs zwischen dem mittleren und späten Teil der späten Kreidezeit in der östlichen Provinz Shandong, China: Beweise aus Dinosaurier-Eierschalen”. Chinesisches Wissenschaftsbulletin. 58 (36): 4663–4669. Bibcode:2013ChSBu..58.4663Z. doi:10.1007 / s11434-013-6059-9. S2CID 131373599.
Recent Comments