ガリミム – ウィキペディア、無料百科事典
ガリミム ( ガリミムス ) – 一種の絶滅した恐竜、オルニトミモサウルスのグループとオルニトミム家のテロポダであり、約7,000万年前の白亜紀後期の現代モンゴルに住んでいます。これはオリニトマイムの最大の代表であり、成人は長さ6 m、高さ1.9 mに達し、重量は約440 kgでした。それは羽で覆われている可能性があり、それは彼の親relativeで確認されました。小さな明るい頭は、横に向かって監督された大きな目を着ていました。他のornition ornitisと比較した負債は、他の種よりも幅が広く丸みがありました。歯は発生せず、それらの不在はケラチンのくちばしに置き換えられました。下顎は繊細でした。多くの椎骨は、内部の空域を示す穴を着ていました。胴体に対する首は長かった。手は、Ornitomimosaursの最短の手は、湾曲した爪のある3本の指で終わりました。前肢は、長い後部と比較して弱いように見えました。
開発のさまざまな段階にある個人の化石は、1960年代にモンゴルのゴビ砂漠でのポーランドモンゴルの古生物学的遠征によって発見されました。この地域からの大きなスケルトンは、新しいタイプと種によって選択されています。 Gallimimus Bullatus 1972年。ジャンルの名前は「鶏を模倣する」を意味し、動物の頸椎と噂(鶏)を結び付ける類似性を指します。種のepithetは、彼の青年期に古代ローマ人が着用したお守りである雄牛に由来しています – 雄牛の構造はガリミマの頭蓋骨の基部にありました。当時の化石 ガリミムス 彼らは、オリニトミマの最もクラスで最も保存された遺跡を構成し、このグループの最も有名な代表者の1人であり続けています。 Ornitomimファミリーは、Ornitomimosaurusのグループに属します。モンゴル出身のモンゴル出身のアンセリミマも最も近い親relativeと見なされていました。
ornithomimeとして ガリミムス 捕食者から逃げることができるスピードのおかげで、彼はハンドルでした。彼が達成した速度は42〜56 km/hと推定されます。彼は、罪人で観察された罪人に匹敵する良い視力と知性を持つことができました。彼は、1つの岩に保存されている数人の個人の発見によって示されているように、グループでの生活につながる可能性があります。ガリミマダイエットやその他のornitiTomimeダイエットについてさまざまな理論が描かれました。首の重要な機動性は、地球上で小さな成果を見つけるのに役立つ可能性がありますが、動物は日和見的な雑食者になる可能性があります。また、ガリミムは水フィルタリングに小さな円柱状のくちばし構造を使用したことも示唆されましたが、これらの構造は、草食生成術を示す硬い野菜食品を食べることの組み合わせでもあります。 ガリミムス これは、彼が親relativeのAnserimimaとDeinocheiraの間に住んでいたNemegt層の最も一般的なornitomimosaurusです。彼は映画に贈られました ジュラシック・パーク 、重要な – 特殊効果の歴史において、シーンは、鳥に似た方法で人気のある見方に従ってそれを示しています。
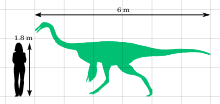
ガリミムス 彼はオリニトミム家の最大の代表者です。成人(標本IgM 100/11)であったホロタイプは、長さ約6 m、腰の高さ1.9 mを測定しました。彼の頭蓋骨は長さ330 mmで、大腿骨は660 mmでした。動物の体重は440 kgと推定されています。比較のために、ZPAL MGD-I/94の若年層の個体は、長さ約2.15 m、腰の0.79 mの測定値であり、重量は約26 kgでした。 [初め] 。化石に密接に関連するオルニトミマに基づいて、ornitomimosaursがふっくらしたことが知られています。大人は、肘のタブの遺跡の存在によって示されるように、構造の翼のようでした。 Lotkiはこの場所のこの場所で上昇しました [2] 。
頭蓋骨 [ 編集 | コードを編集します ]

背骨と比較して、ガリミマの頭は非常に小さくて軽かった。長い銃口のおかげで、ガリミマの頭蓋骨は他のornititomimeよりも長く、口は上部のプロファイルに穏やかに落ちていました。若い人のこの機能はあまり目立ちませんでした。サイドプロファイルは、他のオリニトマイムとガリミマによって区別されていました。また、前半に狭窄が不足しているため、顎の前面にある就学前の骨の下端は水平方向に走るのではなく頭を上げていました。上から見た口は、先端が大きく、丸い(U字型)スケートを取りましたが、北米のオリニトマイムではV字型でした。同じ家族の他の代表者と同様に、大きなアイソケットが側面に向かっていました。目の後ろの頭蓋骨の側面の側頭領域は深く、皮下窓(目のソケットの後ろにある穴の下) – 密接に関連するstrutiomimaによって出会うよりもほぼ三角形で小さくなりました。頭頭の骨に沿った背面の頭蓋骨の丸天井は、筋肉のトレーラーの深い痕跡で飾られていました。薄い壁の中空の骨は、頭蓋骨の底にしわプレートが開いた梨のような球根状の構造を形成しました。内側の鼻孔は大きかった、彼らは口蓋の後ろに横たわっていた。 [3] [4] [5] [6] 。
薄い骨によって構築された繊細な下顎は、正面から細いタイルであり、根に向かって少し深くなりました。彼女の正面は、顎の先端と下顎の間に唇が閉じていても、シャベルを思い起こさせました。現代の灰色のカモメの建設に似ています。誓約の誓約、筋肉の付着の場所は、背中を顎を骨折して動く場所であり、主に角度の骨で構成されていました。一方、顎の最大の骨は、甲状腺機能亢進症の骨でした。これは、テロポードによく見られる特性です。下顎の側面に向けられた下顎の窓は細長く、比較的小さかった。そよ風の虫垂やベニアの骨はありませんでしたが、他の獣脚類とは対照的に、それらの不在は、オルニトミモサウルス、テリジノサウルス、鳥などのくちばしを備えた神経節の間でよく観察される特徴です。頭蓋骨の形状と割合は、動物の成長中に大幅に変化しました。頭蓋骨とアイソケットの背面は相対的なサイズを減らし、口は伸びました。今日のワニには同様の変化が観察されています。また、頭蓋骨は若い人でも比例して大きかった [3] [7] 。顎もあご ガリミムス 彼らは歯を着用しませんでした、彼らの前部はケラチンホーンくちばしでケラチンで覆われていました( Rhamphotheca )。彼は、顎の骨に栄養素がないことによって示されるように、北米の親relativeよりもそれらの小さな部分を占有することができました。くちばしの内面は、適切な性質が依然として議論の対象となっている小さな密に詰め込まれた均等に分布した柱構造によって開発されました。それらの最長は前面にあり、背面が下部と下に横たわっています [5] [7] [8] 。
脊椎 [ 編集 | コードを編集します ]

脊椎 ガリミムス 彼には64〜66個の椎骨、つまり他のオリニトマイムよりも少ない。階段は大手で、平らな前面と凹面の背面がありました。例外は、最初の6つの尾椎骨で、その背面も平らで、両方の平らな表面を備えた尾の尾 – 増幅がありました。茎の多くには胸膜と呼ばれる穴があり、おそらく空気圧骨でした(中空の穴はエアバッグで満たされていました)。首は、頭の後ろに接続する最初の円である桁を除いて、10の頸部、長く、広い椎骨によって支えられていました。アトラスは、その表面として、発見されたものや他のornithomimidaeとは異なっていました セントラム間 彼女は凹面ではなく下向きに寄りかかって前進しました。首は他のオリニトマイムよりも胴体に対して比例して大きいようです。それは2つの別々の部分に分けられました:近位頸椎は、側面図のほぼ三角形の茎によって特徴付けられ、後方に狭くなりました。また、低い椎骨アーチと短い広い関節付属物(Zygapophys)もありました。遠位頸椎には、コイルメッキの茎があり、徐々により長く、薄い関節付属器がありました。椎骨の穴は小さく楕円形でした。椎骨の後部は、背中に最も位置する頸椎をカウントしていない、長くて低く、鋭い櫛を形成して辛い付属物が出てきました [3] [9] 。
海嶺 ガリミムス コイル、短い色合い、より深く、背中に長く、13の背側椎骨を支えました。 rib骨との接続を提供する横方向の付属物、後方に移動するにつれてわずかに長い長さで。最初の2つの背側は深い空気圧穴を着ていましたが、次の2つの穴には浅い穴がありました。有効なとげのある付属物は、三角形または四角形の形状に似ていました。十字架は、長さに等しい5つの十字椎骨によって建設されました。ステップにはコイル、平らな側面、義務があり、一部の標本では深い穴に入りました。とがった付属物は、長方形の形をとり、背側の同等物よりも広く高かった。股関節板の上部海岸の高さまたは等しい高さは互いに分離され、他のornithomimidaeには組み合わされていました。尾は、36から39の椎骨に覆われており、マントのフロントシェードがあり、背面にほぼ三角形の細長い。とがった付属物は、高くて平らで、小さくて小さく、そこで櫛のようなものに入りました。空気圧の唯一の兆候は、辛い付属物と最初の2つの尾部の横断付属物の間の深い穴でした。すべての前椎骨にはrib骨があり、頸部のrib骨は大人にのみ対応する椎骨に関連付けられています [3] [9] 。

手足 [ 編集 | コードを編集します ]
短くて湾曲したヘラは、フロントエンドで薄く、後部は厚かった。上から下まで大きくて厚く、パン粉と比較的接続されていませんでした。一般に、前肢は他のornitionの相同体と有意な違いはありませんでした。この家族のすべての代表者で、彼らは比較的弱かった。上腕骨は、ほぼ丸いセクションで、長くねじれていました。その上部は比較的小さな肩と穴を開けた櫛を着用し、筋肉の付着の小さな表面がフロントエンドを動かすことを保証しました。細長く、長く、ひねられていない肘の骨は、ほぼ三角形のシャフトによって特徴付けられました。大人には大きな肘の付録があり、若い標本ではあまり発達していませんでした。長くて細長いradial骨骨は、それ以上よりわずかに広い端に端を持っていました。手は他のornitomimosaursの手に比べて比例して短かった。また、上腕骨の長さとその長さの代表者間の最も低い関係によって特徴付けられました。さらに、有意な違いはありませんでした。自分に似た3本の指で終わりました。それらの最初は最も強い、つまり親指でした。その後、2番目の指で最も長く続き、3番目は最も弱いものでした。遠位の杭は強く、少し湾曲しており(親指の場合はほとんど)、あらゆる側に深い溝で横方向に平らになりました。これらのステークはお互いを連想させ、3つ目は前のものよりも少し小さかった。成長中、前肢は比例して長くなりました [3] [十] [4] 。
長くて細長い骨盤は、恥骨足で終わりました。これは、オリニトミモサウルスの一般的な特徴であり、前面と背面に拡大しています。後肢は他のオリニトマイムとニコとは異なり、他のテロポードよりも比例して長くなりました。ほとんど単純な大腿骨は長くて細いので、シャフトは横方向に平らになりました。長くて単純な脛骨は、端に2つのよく発達した顆を形成し、端が近く、平らな端がありました。 fi骨は平らで薄く、上端が広く、下部に向かって狭くなっていました。中足骨の3番目の骨の下半分は広く、両側に隣接する中足骨骨で部分的に覆われていましたが、長さの半分が激しく狭まり、それらの間を切り開いていました。 3番目の指は、他のOrnithomimidaeよりも肢全体に対して短かった。彼らと同様に、つま先はありませんでした。遠位杭は、側面の下部で平らになり、2つは外側がわずかに外側に傾いていました。骨の相対的な長さは、動物の成長とともに非常にわずかに変化しました [3] [4] 。

1963年から1965年の間に、ポーランド科学アカデミーとモンゴル科学アカデミーは、モンゴルのゴビ砂漠へのポーランドとホンゴの古生物学的探検を組織しました。ネメグト渓谷の砂浜の堆積物で発見された恐竜の遺跡には、ネメグト、ツァガン・クシュウ、アルタン・ウラIV、ナルニア・ブラックのさまざまな成長段階にあるオリニトマイムの多くの遺跡が含まれています。 3つの部分的に完全なスケルトンが見つかりました。2つは頭蓋骨と多くの断片的な遺跡です。それらの最大(後でホロタイプとして認識されました Gallimimus Bullatus )1964年にツァガン・クシュウのポーランドの古生物学者のゾフィア・キエラン・ヤボロウスカを発見しました。動物は、骨盤の下に頭蓋骨を置いて、背中に横たわっている位置に保存されました。同じ年に、Nemegtの小さな標本も見つかりました。 1967年に、ネメグト渓谷の外のBgeen Tsavでのモンゴル遠征によって、前肢のない小さな骨格が発見されました。化石はモンゴル人、ポーランド人、ソビエト/ロシア科学アカデミーを所有しています [3] [11] [12番目] 。 1968年にワルシャワでいくつかのスケルトンが展示され、当時は強制的に強化された態度で組み立てられました。現在、これらの恐竜がより水平な態度を採用したと想定されています [13] 。

1972年、古生物学者HalszkaOsmólska、Ewa Roniewicz、Rinchen Barsboldが新しいジェネリックとジャンルの名前を調理しました Gallimimus Bullatus 、Tsaagan Khushuuを使用した最大の収集された標本IgM 100/11に基づいていました。これは以前はG.I.No.dps 100/11およびMPD 100/11として知られていました。ジャンルの名前は、ラテン語から来ています ガルス 鶏肉とギリシャ語から 甘やき フォロワーを意味します。彼らはここで首の椎骨の前面を紹介しました。これは、鶏で観察されたことを連想させます。種のepithetはラテン語から来て、そのソースを取得します 雄牛 、彼の首に古代ローマが着ている金色のカプセル。彼女はそれを、頭蓋骨の基部にある横隔膜上の球根状のカプセルで研究者と結び付けました。そのような構造は、爬虫類では以前に説明されていなかったため、珍しいように思えました。ホロタイプには、歪んだ口、不完全な顎、椎骨シリーズ、骨盤、および手と足の骨がなくなったほぼ完全な骨格が含まれていました [3] [14] 。若い人は他の部分的に完全な骨格を残しました。ツァガン・クシュウを添えたZPAL MGD-I/1には、先端が欠けている頭蓋骨、破損した椎骨、断片的なrib骨、上肢の縁と前足、および不完全な左後肢がありました。 ZPAL MGD-I/94では、ネメグトには頭蓋骨、ガールフレンド、テールチップ、上肢の縁、同じ手足自体がありませんでした。最小の標本IgM 100/10は、Bgeen Tsavで発見され、上腕、前肢、いくつかの椎骨とrib骨がいませんでした。 Osmólskaと同僚は、合計25のよく知られている標本を交換しました。そのうち9つは個々の骨です [3] [十] 。
化石のタイプの説明中 ガリミムス それらは最も完全で最高の保存された化石材料であり、このグループの最も有名なメンバーの間では常にタイプが残っています。 Ornithomimidaeは以前は主に北米大陸から知られていましたが、アジアからのみ出会っただけでした Archaeornithomimus 、そして彼の場合、頭蓋骨は以前には見つかりませんでした。最初の発見以来、より多くの標本がモンゴル人によって管理されている国際遠征を見つけました [3] [十] [15] [13] 。 3人のスケルトンガリミマ(ホロタイプを含む)は、モンゴルの恐竜の化石の巡回展の一部となり、多くの国を横断しています [16] [17] 。化石を奪うことは、21世紀のモンゴルで重要な問題となっています。いくつかのガリミマ標本も盗難の犠牲になりました。 2017年、Hang-Jae Leeと同僚は、2009年にスタックフットとともに発見された化石化された痕跡を報告しました ガリミムス (MPC-D100F/17標本)。残りのスケルトンは、他のいくつかの標本とともに、早めに略奪されているように見えました ガリミムス 、発掘、廃棄物、骨折した骨が採石場の周りに散らばった後の空の場所で示されました。それにもかかわらず、発見されたものは、化石化された痕跡と動物の化石化された体とともに珍しいものでした。痕跡のいくつかはornitomimeの足に合っていますが、他の痕跡は異なる種類の恐竜を残しています [18] [19] 。

Osmólskaと同僚が合格しました ガリミムス 1972年のOrnitomim科(Ornithomimidae)に、彼の最も近い親relative、北米と一緒に struthiomimus 、両方のタイプと比較して困難を後悔している。なぜなら、他の鳥類はその時点で保存されていない遺跡のために知られていたか、適切に記述されていなかったため [3] 。 1975年、キーラン・ヤワロウスカは、多くのアジアの恐竜が北米の親sと同じ家族に配置されたが、このランクは現代の鳥の場合よりも包括的であるように見えると述べた。彼女はそれを強調した ガリミムス 彼はガチョウやアヒルに似た丸い梁を持っていました。北米大陸の鳥類はくちばしを引き締めました。現代の鳥の場合のこのような違いは、それらを2つの別々の家族に置くことになります [15] 。 1976年、バースボールドはオリニトミモサウリ(オリトミモサウリア)の新しいグループにオルニトミミダエを配置しました。 2003年、小林とジュンチャンリュはそれを発見しました anserimimus 彼はガリミマの姉妹分類群でした。一緒に、彼らは2006年に小林とバーズボールドによって確認された北米タイプの進化レールを形成しました [十] [5]
1988年、古生物学者のグレゴリー・S・ポールは、オリニトミムの頭蓋骨のより大きな類似性を以前考えていたよりも自分自身に示したため、すべての種を1つのタイプに移しました。 ornithomem 、ガリミマを作る O. Bullatus [初め] 。しかし、2010年には、彼は「 ガリミムス ( struthiomimus )) Bullatus 「、2016年に彼はジャンル名のみを使用しました ガリミムス [20] [21] 。他の著者は通常、種を別々のタイプに配置しました [4] 。 1988年、非公式に定義された「サンチャウス」のオリニトミマサークルが報告されましたが、属に割り当てられました ガリミムス 1990年の古生物学者ドン・ジミングと同僚による不確実な種(sp。) [14] 。 Barsboldは、ほぼ完全なIGM 100/14スケルトンを「Gallimimus Mongoliensis」と非公式に言及していましたが、 ガリミムス 吉村小林とバースボールドの建設のいくつかの詳細で、彼らは2006年に別のタイプに配置することを提案しました。 2000年、古生物学者のフィリップ・J・カリーはそれを提案しました anserimimus 、モンゴルの単一のスケルトンからのみ知られている、若い同義語です ガリミムス 、その結果、KobayashiとBarsboldの批判と会う。彼はタイプのいくつかの違いを指摘した。 Barsboldは、新しいGallimima標本の間でいくつかの形態学的なバリエーションに気づいたが、彼はそれを公開しなかった [十] 。
Li Xuおよびその他の2011年によると、Ornitomimesの間のGallimimaの位置は、次のクラドグラムを示しています [22]

Ornithomimosauriaは、ターゲットに含まれるManiraptoriformesに含まれており、鳥を含むテロポードのグループも含まれています。初期のornitomimosaursは歯を持っていて、グループのより進化的な代表者によって失われました [23] 。 2004年、マコヴィッキー、小林、カリーは、オルニトミモサウルスの初期の進化の歴史のほとんどがアジアで走ったことを示唆しました。しかし、彼らはオリジナルの存在を認めました ペレカニミムス ヨーロッパでは、分類を複雑にします。グループは、アジアからベリンギアを介して北米大陸に1回または2回広まらなければなりませんでした。恐竜の他のグループと同様に、オルニトミモサウルスは、ヨーロッパがアジアからアジアから分離した後、大部分がアジアと北米に限定されていました。 [4] 。
1994年、古生物学者のトーマス・R・ホルツは、横隔膜上に球根状構造の存在などの特徴に基づいて、ornitomimosaursとtroodontsをグループ化しました。彼は、種の種に基づいて、提案されたBullatosauria Clampを呼びました Gallimimus Bullatus 、この解剖学的特徴についても言及しています [24] 。しかし、1998年、ホルツはトルドンツが基礎マニラプトルであるという結論に達しました。これは、この後者のグループのすべてのメンバーがBullatosauriaに属し、このグループManiraptoriformesの若い同義語を作ることを意味します。したがって、名前は使用されませんでした [25] [26] 。

頸椎 ガリミムス 彼らは傾斜した頭を示し、35度の角度で落ちます。 Osmólskaと同僚は、Gallimimの手がつかむことができないことを発見しました。恐竜は残りの指で親指に反対することができませんでした。彼らはまた、手が手に比べて弱いことを示唆しました。たとえば、別のornitomimosaurus Deinocheirus 。彼らは、1972年から古生物学者のダル・ラッセルによって提案されたオリニトメモウの生物学の解釈に同意し、罪のない鳥の現代の鳥よりも機敏ではありませんが、動物の手を見ました。彼らの捕食者はカラフルなビジョンを持っている可能性があるため、彼はこれがオリニトマイムの色に影響を与える可能性があることを示唆しました。 [3] [27] 。 1982年、古生物学者のリチャードA.シュルボーンは、実行の速度を推定しました ガリミムス 42-56 km/hで。彼は、Ornitommsが今日のダチョウの速度と一致していないことを発見しました。部分的には上肢と尾の増加に関連して、70〜80 km/hに達します。 [28] 。

1988年、パウロは、オリニトマイムがアイソケットに最小の可動性を備えた眼球を平らにしたことを示唆したため、頭の動きが大きな役割を果たすと示唆しました。彼らの目は治療の鳥に似た他のいくつかよりも横向きになったので、双眼視の領域は非常に限られていた。このような適応は、動物が脅迫的な捕食者を探すのに役立ちます。パウロは、比較的短い尾を認識し、体重を減らし、スピードへのオリニトミムの適応のためのつま先の欠如を認識しました。彼は、これらの恐竜が自分自身を擁護し、嘆き、蹴ったことを示唆したが、最大限の程度まで彼らは素早い脱出で構成されていた [初め] 。 2015年、渡辺akinobuと他の人はそれを発見しました – まさに Deinocheirus 私 Archaeornithomimus – ガリミムス 彼はornitomimosaursの中に空気圧の骨格を持っていました。空気圧骨は、現代の飛行の鳥にとって簡単になりますが、未知の恐竜を伴う機能は不明のままです。それは、より大きな骨の質量を減らすこと、高い代謝率との関係、運動中のバランスまたは体温調節について提案されました [9] 。
2017年、LeeとのLeeは、2009年からの発見の痕跡とともにGallimimaの足がどのように振る舞ったかを説明するために、さまざまなタタゴノミック条件を提案しました。化石は痕跡で死んだ動物から来る可能性がありますが、泥の中の足の深さは浅すぎるかもしれません。ザウロップサイドは洪水を殺す可能性があり、その後彼の死体は池に埋葬されました。しかし、シルトと砂の層は洪水を示すのではなく、乾燥した生息地を示しており、化石の周りのひび割れた堆積物は、この場所にいるときに動物がまだ生きていたことを意味します。したがって、著者らは、痕跡が乾燥に先行する別の動物の死期の前に長い間残され、おそらくそれらのどれも残されていないことを示唆した。恐竜は池の底を歩いて、集落の周りを歩き回ることができました。そこでは痕跡がすでに刻印されており、彼はそこで亡くなり、泥の中に沈みました。足は、腱が曲がったときに、体の崩壊中、そしてその後の重い恐竜が発生したときに、骨格の残りの部分から立ち往生し、切断することができました。この地域は、干ばつや飢erのために、おそらくガリミムの大量死を表す単一の墓地(可能な数の個人に基づいて)である可能性があります。動物は同時に死んでいるようです(作られた骨が層序的に同一である後の空の空洞)は、群れのライフスタイルを示している可能性があります ガリミムス 、他のornitometsの場合にも提案されています [19] 。
発達 [ 編集 | コードを編集します ]

生物学者のローマン・ポーリッキーとP.ボレチャワによる1987年の研究では、さまざまなガリミマ標本の骨におけるカルシウムとリンの年齢関連の違いが示されました。彼らの態度は、若いまたは中間の個人で最も高く、年齢とともに減少しました [29] 。 1991年、研究者たちは、古い人の骨が最も多くの鉛と鉄を含んでいたと報告しました。 [30] 。 2000年に生物学者ジョン・M・レンズバーガーとマヒト・ワタベによって発行されたさまざまな恐竜の骨組織学の検査により、骨チャネルとコラーゲン繊維が明らかになりました ガリミムス また、他のオルニチトミメンは、哺乳類のホモロジストとは対照的に、哺乳類の構造に似ているのではなく、哺乳類よりも鳥で観察されたものに似ていました。これらの違いは、骨形成のプロセスと速度に関連する可能性があります [最初に30] 。 2012年、古生物学者のDarla K. Zelenityskyと同僚は、成人のOrnitomimosaursが上肢に翼のある構造を持っていて、若い人はそれらを持っていなかったという結論に達しました(標本によって証明されているように ornithomem )、これらの構造は、求愛、交尾ショー、蒸気で役割を果たすことができる高等の性的特徴を構成します [2] 。
食べ物 [ 編集 | コードを編集します ]

オスモルスカと同僚は、ガリミマの首の前面のかなりの移動性(首の後ろは硬い)に気づき、この地域の椎骨のアーチは、鶏やその他のキュアリーで見られることに似ており、同様の食物習慣を示しています。彼らはそれを発見しました ガリミムス 彼はアヒルやガチョウに似ていたので、スタガーは小さな生きた獲物を食べました。首の運動性は、目が頭蓋骨の側面に配置されていたため、地面に獲物を見つけるときに有用でした。研究者は、すべてのオリニトマイムの同様の栄養習慣を採用しています。彼らは、ラッセルがオルニトミミダエのくちばしを殺虫性の鳥と比較したことに気づきました。 Osmólskaなどは彼らの能力を提案しました ガリミムス 頭蓋骨の後ろにある骨間の目に見えるゆるいつながりに関連するキネシスの場合。この機能により、個々の骨が互いに比較的移動できます。研究者はまた、恐竜が口に食べ物を供給するために彼の短い前肢を使わず、略奪して地面を掘ってそれを手に入れることを提案しました [3] 。手 ガリミムス 1985年に古生物学者が1985年に書いたように、彼らはそれらを発作とつかむためにむしろ使用しているStrutiomimの腕に道を譲ることができました。 [32] 。
1988年、パウロは、以前に示唆されたように、彼らが小動物と卵、植物を食べたことに従って、オルニトマイムの雑化の見解に反対しました。研究者は、ダチョウとEMUが主に植生を食べていることに気づき、オリニトマイムの頭蓋骨は、腸の内容によって証明されるように、小枝を壊すのに十分強く、絶滅したMOAの頭蓋骨のほとんどに似ていたことに気付きました。彼はさらに、Ornithomimidaeが硬い植物によく適応し、手を使って枝を口の領域に曲げることを示唆しました [初め] 。古生物学者ヨルン・フルムは2001年に顎と下顎の同様の構造に関連して示唆した ガリミムス 彼はカモメのような日和見的な雑食者だったかもしれません。研究者は、顎の前面と部分の間の動きに対して顎の中にタイトなジョイントを固定していることに気づきました [7] 。

2001年、古生物学者のマーク・ノレル、マコヴィッキー、カリーは頭蓋骨について説明しました ガリミムス IGM 100/1133およびSkull ornithomem 保存された軟部組織残留物がくちばしで。ガリミマのくちばしの内側は、柱の構造で装飾されていました。著者は、食物の操作、堆積物を排出し、食物粒子を排出し、食物粒子をろ過するために使用され、給餌中に植物を遮断するために使用されるブリキの弓のプラークに似ていました。彼らは、植物、軟体動物、配偶者、穴を食べる普通の平らなものが、最も可能性の高いガリミマ構造を持つ構造を持つ現代の缶であることを発見しました。著者は、Ornitommsがおそらく大型動物を狩るためにくちばしを使用していないことに気づいています。また、移行環境でも一般的であり、乾燥しています。これは、食物の水源への依存を示唆しています [8] 。これが正しい解釈である場合、 ガリミムス 最大の既知の土地フィルターになります [4] 。
2005年、古生物学者のポール・バレットは、鳥の食物をろ過するような毛が柔軟になっているように見えることに気づきました(お互いの証拠がないため)。代わりに、それらはより薄く、等しい間隔で、カメやハドロサウリーのくちばしの垂直の折り目でした。これらの動物では、これらの尾根は草食動物の食事に関連しており、硬い植生を切るために使用されます。バレットは、ガリミマの弓の尾根はくちばしの内面の自然な鋳造であり、これは非常に繊維状の食物の草食生成と消費を示すことを示唆しました。オリニトマイムの遺跡で多くの胃腸岩の発見は、筋肉の胃の存在を示しており、これらの石は適切なマウスピースを持たない食品を粉砕するために使用されるため、草本を示唆しています。バレットは、ガリミマを含むOrnitomimosaursが採取した食物の量を推定しようとしました。動物の体重として、彼は440 kgを採用しました。エネルギーの需要は、特定の仮定の採用を必要とし、科学者はいくつかの個別の推定を行いました。 Gallimimに、現代の哺乳類を特徴付ける迅速な代謝を割り当てることにより、1日あたり29 MJを超えることができます。同様の結果は、今日の鳥に典型的な代謝のペースを採用した結果です(ただし、エネルギー需要のほぼ2倍の小さなスズメ鳥ではありません)。ガリミムがトカゲで観察されたトカゲに近い代謝を持っていた場合、この問題は、冷たい血液(約1.5)は言うまでもなく、温かい血色のもの(1日9 mj未満)でさえも完全に異なって見えます。このような大きな矛盾は、毎日吸収される食物の推定質量が大きく異なることを意味します。最も低い結果である1日70 gは、冷血トカゲの代謝のペースと栄養価の高いリング、甲殻類、またはシウニプロドウィ軟体動物で作られたメニューを仮定することで得られます。最高の結果である3 µkgを超えると、他の軟体動物を食べるスズメ鳥の代謝とガリミが服用されます。これは、日中に数千リットルの水を撮影することを意味しますが、それはバレットトウェイが実現可能ではないと思われます。フィルターとして、彼はそのような量の食物を受け入れることができません。研究者はまた、Ornitommsが移行中の生息地だけでなく、フィルターを維持するのに十分な量の水を確保しないように乾燥していることを発見しました。 [6] 。 2007年、古生物学者のエスパンM.クヌッツェンは、現代の鳥で観察された現代の鳥と比較して、オルニトマイムのくちばしの形状を書きました。 [33] 。

ガリミムス 彼は、モンゴル南部のゴビ砂漠でのネメグト層で知られています。このフォーメーションの放射測定の年代測定は決して実行されていませんが、その岩で見つかった動物相は、約7,000万年前のクレダの早期メストリックを示しています。 [34] [35] [36] 。典型的な場所の集落 ガリミムス ツァガン・クシュウには、ラバ、泥、砂、そしてあまり一般的ではない砂岩の薄い層が含まれています [12番目] 。揺れ施設は、過去に川の水路の存在、泥だらけの平原、浅い湖、flood濫原が環境を形成することを示唆しています。 [37] 。大きな川の水路と塩の堆積物は、古いバルン・ゴヨット層やダジャドック層に関連するものよりもはるかに湿った気候であることが証明されていますが、カリッシュの集落は周期的な干ばつを示しています [38] 。このNemegt Valleyの化石の可能な骨 ガリミムス 、それらは周辺地域の化石よりも放射性が高い。これは、地下水の浸漬のおかげで骨に蓄積されたウランに責任があるかもしれません [13] [39] 。
Nemegt Riversは、幅広い生物の家でした。軟体動物と他のさまざまな水生動物がそこで発見されました:魚、カメ [34] 、クロディロモルフ、その中 Shamosuchus [40] 。哺乳類はこの層の岩ではまれですが、エナンチオールには多くの鳥が見つかりました ビート 、hesperornithes ジュディノリス 私 Teviornis 、おそらくブリキ。草食の恐竜は、アンキロサウルスを代表しています タルチア 、Pachycephalosaurus Prencefal、大きなハドロザウルドZaurolof i Barsboldia 、 結局 [34] [41] 。ガリミムの隣に住んでいる述語テロポード バガラタン 、Troodonts Borogowia、Tochizaur i ザナナバザール 。草食性または雑食性のテロポードには、テリジノサウルスなどのテリジノサウルスや、エルミザウルのようなovraptorosaursが含まれます。 非gtomaia 私 Rinchenia [42] 。他のornitomimosaursの遺跡がそこに発見されました、 anserimimus 私 Deinocheirus 、しかし、このガリミムはNemegtのこのグループの最も一般的な代表です [十] 。

ガリミムス 1993年に映画で発表されました ジュラシック・パーク スティーブン・スピルバーグ監督。元の小説の同様のシーンは、ハドロサウリーを描いた。スピルバーグは急いでいる群れのシーンを望んでいましたが、彼はそれを達成する方法を知りませんでした。当初、時間を経てそれを実現しようと試みられました。当時、コンピューターのアニメーションを備えた動物のイメージングに小さな信仰が置かれていましたが、特殊効果を担当する産業用光と魔法の会社は、可能性を探求するためのメーカーの同意を得ました。彼女はガリミマスケルトンのコンピューターモデルを作成し、スケルトンの群れのコースを描いたテストアニメーションを実行し、後にガリミムの完全に生成された群れを追いかけるティラノサウルスを実行しました。制作チームは、これまでに似たものがなかったため、これを非常に熱意で受け入れました。 Spielbergは、映画にシーンを含め、Time -lapse -Animationではなく恐竜の他のショットでコンピューターグラフィックを使用するために発見しました。 [43] [44] 。 ガリミムス ダチョウのショットとガゼルのhensingの分析のおかげで彼らは提示されました [45] 。キーラン・ヤワロウカは美しいシーンを呼んだ [13] 。フィルム恐竜は、映画で最も一般的に示されているCGIアプリケーションの1つになりました。彼らは以前の特殊効果よりも「生きている」ことが認識されていました [46] 。
ジュラシック・パーク 彼は鳥の群れの行動を強調した ガリミムス 、初期の鳥の親relativeを代表しています。映画の群れ全体が一緒になって、小さな個人がグループの中央にとどまり、より大きなものによって保護されていました [43] 。このシーンでは、映画の古生物学者であるアラン・グラントのヒーローは、捕食者から逃げる鳥の群れと同じように走る方向の変化に気づき、彼が同じように鳥を見ることは決してないという速くて優雅なガリミムの走りを見ている対談者に追加します。このイメージは、メディアで恐竜を提示する伝統的な方法とは対照的です。この映画は、恐竜の社会的認識を変えるのに役立ちました。このシーンや他のシーンは、科学コンサルタントのペレオンティストJacek Hornerのおかげで紹介された鳥の進化に関する見解を、まだ物議を醸す撮影中に新しいシーンを反映しています。 [47] [48] [49] 。にもかかわらず ガリミムス そして、他の恐竜は羽のない映画に贈られました。その部分は、このグループの動物の中でどのように広まっているかがまだ知られていなかったからです。 [47] [50] 。
オーストラリアのクイーンズランドにあるラーク採石場(世界最大の恐竜の痕跡の密度がある場所の1つ)の痕跡は、ガリミムとのシーンのインスピレーションと科学的支援として機能したと主張されました。これらの痕跡は、最初にテロポッド群からの捕食者の外観によってownされた恐竜のパニックの残留物として解釈されました。このアイデアは批判されました、とされるテロポッドの疑いは、実際にはムタバザウラと同様に、別のグループの草食動物である可能性があります。コンサルタント ジュラシック・パーク 彼は、痕跡が本当に映画のインスピレーションであることを否定しました [51] [52] [53] 。
2000年3月24日に発行されたPoczta Polska、シリーズのGallimimaを示す郵便印刷 先史時代の動物 – 恐竜 0.7 Zlotysの宗派について。スタンプのプロジェクトの著者は、Jacek Brodowskiでした。スタンプは、2,400,000 PCの循環で、蛍光紙にオフセット技術を使用して印刷されました。 [54] 。動物はスタンプに記載されています ガリミムス [55] 。
- ↑ a b c d G.S. ポール 、 世界の略奪的な恐竜 、ニューヨーク:Simon&Schuster、1988、s。 384–386、393–394、ISBN 978-0-671-61946-6 。
- ↑ a b D.K. ゼレニツキー 私はイニ 、 北米からの羽のない非鳥の恐竜は、翼の起源に関する洞察を提供します 、「科学」、338(6106)、2012年、s。 510–514、doi: 10.1126/science.1225376 、PMID: 23112330 、bibcode: 2012Sci … 338..510Z 。
- ↑ a b c d そうです f g h 私 j k l m H. オスモルスカ 、 と。 Roniewicz 、 R. Barsbold 、 新しい恐竜、 ” Gallimimus Bullatus ” n。 Gen。、n。 sp。モンゴルの白亜紀上部から(ornithomimidae) 、「Palaeontology Polonica」、27、1972、pp。103–143 。
- ↑ a b c d そうです f P.J. マコビッキー 、 と。 小林 、 P.J. カリー 、 ornithomimosauria 、 [の:] デビッド weishampel 、 ピーター ドドソン 、 halszka オスモルスカ 、 恐竜 、WYD。 2、バークレー:カリフォルニア大学出版局、2004年、s。 137–150、ISBN 978-0-520-24209-8 。
- ↑ a b c と。 小林 、 J.-C. ルイ 、 中国の白亜紀後期からの群れの習慣を持つ新しいornithomimid恐竜 「Acta Palaeontologica Polish」、48(2)、2003、s。 235-259 ( 。 )) 。
- ↑ a b 午後 バレット 、 ダチョウの恐竜の食事(セロポダ:ornihomimosauria) 、「古生物学」、2、48、2005、s。 347–358、doi: 10.1111/j.1475-4983.2005.00448.x 。
- ↑ a b c J. ハム 、 「gallimimus bullatus」の下顎 、 [の:] 考え 私はイニ 、 中生代の脊椎動物の生命 、ブルーミントン:インディアナ大学出版局、2001年、s。 34–41、ISBN 978-0253339072 。
- ↑ a b M.A. ノレル 、 P.J. マコビッキー 、 P.J. カリー 、 ダチョウの恐竜のくちばし 、「自然」、412(6850)、2001年、s。 873–874、doi: 10.1038/35091139 、PMID: 11528466 。
- ↑ a b c A. 渡辺 私はイニ 、 Ornithomimosaur ” Archaeornithomimus ”(恐竜:Theropoda)の椎骨空気圧症の椎骨像のイメージングとオルニトミモサウリアの軸の空気圧の再評価によって明らかになった 、「Plos One」、12、10、2015、E0145168、doi: 10.1371/journal.pone.0145168 、PMID: 26682888 、PMCID: PMC4684312 、bibcode: 2015Ploso..1045168W 。
- ↑ a b c d そうです f と。 小林 、 R. Barsbold 、 モンゴルのネメグト層からのornithomimids 、「韓国古生物学会のジャーナル」、1、22、2006、s。 195–207 。
- ↑ と。 キーラン・ヤボロウカ 、 N. ドブチン 、 ポーランド – モンゴルの古生物学的遠征の物語1963-1965 、「Palaeontologica Polonica」、1968、s。 7–30 [アクセス2019-04-14] [アーカイブ 住所 2020-07-12] 。
- ↑ a b R. グレードジアン 、 J. Kazmierczak 、 J. レフェルド 、 ポーランド – モンゴルの古生物学の遠征からの地理的および地質データ 、「Palaeontologica Polonica」、19、1969、s。 33–82 。
- ↑ a b c d と。 キーラン・ヤボロウカ 、 初期の哺乳類の追求 、ブルーミントン:インディアナ大学出版局、2013年(過去の生活)、s。 47–48、50–51、ISBN 978-0-253-00824-4 。
- ↑ a b D.F. glut 、 恐竜:百科事典 、ジェファーソン:McFarland&Co、1997、s。 429–432、ISBN 978-0-89950-917-4 。
- ↑ a b と。 キーラン・ヤボロウカ 、 ゴビ砂漠の後期白亜紀哺乳類と恐竜:1963年から71年のポーランド – モンゴルの古生物学的探検によって発掘された化石は、原始哺乳類と恐竜、アジアと北米の間の動物相交換に新しい光を投げかけます 、「アメリカの科学者」、2、63、1975、s。 150–159、bibcode: 1975amsci..63..150k 。
- ↑ 白亜紀モンゴルの恐竜 、dinocasts.com [アーカイブ 住所 2015-02-02] 。
- ↑ S.G. 西 、 S. ハンセン 、 T. 西 、 恐竜、卵、子供 – 行方不明の世界に関する特別展 、「Geological NYT」、6、1997、pp。24-25 [アクセス2018-05-13] [アーカイブ 住所 2018-05-14] ( デュディ。 )) 。
- ↑ F. 歩兵 私はイニ 、 モンゴルの密猟された恐竜の化石を本国に送還するためのツールとしての地球化学的指紋 、「古地理学、古眼科、古生物学」、494、2017、s。 51–64、doi: 10.1016/j.palaeo.2017.10.032 、bibcode: 2018ppp … 494 … 51f 。
- ↑ a b H.-J. リー 私はイニ 、 モンゴルのネメグト層からの「ガリミムス」フットスケルトンに関連する獣脚類トラックウェイ 、「古地理学、古眼科、古生物学」、494、2018、s。 160–167、doi: 10.1016/j.palaeo.2017.10.020 、bibcode: 2018ppp … 494..160l 。
- ↑ G.S. ポール 、 恐竜へのプリンストンフィールドガイド 、WYD。 2、プリンストン:プリンストン大学出版局、2016年、s。 131、ISBN 978-0-691-16766-4 。
- ↑ G.S. ポール 、 恐竜へのプリンストンフィールドガイド 、WYD。 1、プリンストン:プリンストン大学出版局、2010年、s。 112–114、ISBN 978-0-691-13720-9 。
- ↑ L. コイン 私はイニ 、 中国の河南省にある白亜紀後期のキウパ層からの北米の親和性を持つ新しいオルニトミミッド恐竜 、「白亜紀研究」、2、32、2011、s。 213、doi: 10.1016/j.cretres.2010.12.004 。
- ↑ C. Hendrickx 、 S.A. ハートマン 、 O. マテウス 、 非鳥類の獣脚類の発見と分類の概要 、「脊椎動物の古生物学のパラックのジャーナル」、1、12、2015、s。 1–73 [アクセス2019-04-14] [アーカイブ 住所 2018-06-22] 。
- ↑ トーマスR. ホルツ 、 Tyrannosauridaeの系統発生的位置:獣脚類体系への影響 、「Journal of Paleontology」、68(5)、1994年9月、pp。1100–1117 、jstor: 1306180 。
- ↑ トーマスR. ホルツ 、 肉食性恐竜の新しい系統発生 、「ガイア」、15、1998、s。 5–61 、ISSN 0871-5424 。
- ↑ D. ナイッシュ 、 D.M. マーティル 、 と。 フレイ 、 ブラジルのサンタナ層(?アルビアン、初期の白亜紀)からの新しい獣脚類を含む恐竜の生態学、系統学、および生物地理学的関係 、「歴史的生物学」、16(2–4)、2004年、s。 57–70、doi: 10.1080/08912960410001674200 。
- ↑ D.A. ラッセル 、 カナダ西部の白亜紀後期のダチョウの恐竜 、「Canadian Journal of Earth Sciences」、9(4)、1972、s。 375–402、doi: 10.1139/e72-031 、bibcode: 1972cajes … 9..375r 。
- ↑ R.A. Thulborn 、 恐竜の速度と歩行 、「古地理学、古眼科、古生態学」、38(3–4)、1982年、s。 227–256、doi: 10.1016/0031-0182(82)90005-0 、bibcode: 1982ppp …. 38..227t 。
- ↑ R. Pawlicki 、 P. ソファー 、 化石恐竜の骨のX線微量分析:「Gallimimus Bullatus」骨のカルシウムとリン含有量の年齢差 、「Folia histochemica et cytobiogica」、3–4、25、1987、s。 241–244、PMID: 3450541 。
- ↑ R. Pawlicki 、 P. ソファー 、 化石恐竜の骨のX線微量分析:鉛、鉄、マグネシウムの含有量の年齢差 、「folia histochemica et cytobiogica」、2、29、1991、s。 81–83、PMID: 1804726 。
- ↑ J.M. レンズバーガー 、 M. ワタベ 、 恐竜、鳥、哺乳類の骨の細かい構造 、「自然」、406(6796)、2000、s。 619–622、doi: 10.1038/35020550 、PMID: 10949300 。
- ↑ E.L. ニコルズ 、 A.P. ラッセル 、 「struthiomimus altus」の胸溝と前肢の構造と機能(Theropoda:ornithomimidae) 、「古生物学」、28(4)、1985、s。 64–677 ( 。 )) 。
- ↑ E.K. マドセン 、 存在する鳥のくちばしの形態は、オルニトミイドのくちばしの形態に影響を与える 、「数学と自然科学の学部 – 論文」、2007、pp。1–21 ( 。 )) 。
- ↑ a b c T. Jerzykiewicz 、 D.A. ラッセル 、 中生代後期層序とゴビ盆地の脊椎動物 、「白亜紀研究」、12(4)、1991、s。 345–377、doi: 10.1016/0195-6671(91)90015-5 。
- ↑ R.M. サリバン 、 パキセファロサウリド科の分類学的レビュー(恐竜:Ornithischia) 、 [の:] スペンサーG. ルーカス 、 ロバートM. サリバン 、 西部の内部からの後期白亜紀脊椎動物 、t。 35、ニューメキシコ自然史博物館科学紀要、2006年、s。 347–366 [アクセス2019-04-14] [アーカイブ 住所 2007-09-27] 。
- ↑ F.M. グラッドシュタイン 、 J.G. ogg 、 A.G. スミス 、 地質学的時間スケール2004 、ケンブリッジ:ケンブリッジ大学出版局、2005年、s。 344–371、doi: 10.1017/cbo9780511536045 、ISBN 978-0-521-78142-8 。
- ↑ トーマスR. ホルツ 、 古生物学:恐ろしい手の謎が解決しました 、「自然」、515(7526)、2014年、s。 203–205、doi: 10.1038/nature13930 、PMID: 25337885 、bibcode: 2014natur.515..203H 。
- ↑ M. Novacek 、 燃えるような崖の恐竜 、ニューヨーク:Anchor、1996、s。 133、ISBN 978-0-385-47775-8 。
- ↑ と。 ジョワリアン 、 J. ペンシューズ 、 モンゴルからの異常に放射性の化石骨 、「自然」、214(5084)、1967、s。 161–163、doi: 10.1038/214161a0 、bibcode: 1967natur.214..161j 。
- ↑ M.B. エフィモフ 、 モンゴルの化石ワニの修正 、「sovmestnaya sovetsko-mongol’skaya paleontologicheeskaya ekspedititsiya trudy」、24、1983、S。 76–96 ( ロス。 )) 。
- ↑ J.H. ハム 、 K. 安息日 、 アジアと北米の巨大な獣脚類恐竜:「ターボサウルスバタール」と「ティラノサウルスレックス」の頭蓋骨と比較してください 「Acta Palaeontologica Polish」、48(2)、2003、s。 188 。
- ↑ トーマスR. ホルツ 、 ティラノサウルド 、 [の:] デビッド・B weishampel 、 ピーター ドドソン 、 halszka オスモルスカ 、 恐竜 、WYD。 2、バークレー:カリフォルニア大学出版局、2004年、s。 124、ISBN 978-0-520-24209-8 。
- ↑ a b D. シェイ 、 J. ダンカン 、 ジュラシックパークの製造 、ニューヨーク:Boxtree Ltd、1993、s。 48–52、135–136、ISBN 978-1-85283-774-7 。
- ↑ M. クリトン 、 ジュラシック・パーク 、ロンドン:ランダムセンチュリーグループ、1990年、s。 137–138、ISBN 978-0-394-58816-2 。
- ↑ R. ベアード 、 動物化「ジュラシックパーク」「恐竜」:脅威シーンでの大ヒットスキーマと異文化の認知 、「Cinema Journal」、4、37、1998、s。 82–103、doi: 10,2307/1225728 、jstor: 1225728 。
- ↑ S. 王子 、 真の嘘:知覚的リアリズム、デジタルイメージ、映画理論 、「フィルムクォータリー」、3、49、1996、s。 27–37、doi: 10,2307/1213468 、jstor: 1213468 。
- ↑ a b S. モンタナリ 、 「ジュラシックパーク」が出てきてから恐竜について学んだこと 、2015年 [アクセス2018-05-07] 。
- ↑ D.A. カービー 、 ハリウッドのラボコート:科学、科学者、映画 、マサチューセッツ州:Press、2011、p。127–129、ISBN 978-0-262-29486-7 。
- ↑ D.A. カービー 、 科学コンサルタント、架空の映画、科学的実践 、「科学の社会科」、2、33、2003、s。 231–268 。
- ↑ 私。 フィリップス 、 どのように「ジュラシックワールド」恐竜は実生活を見ていました 、Business Insider、2015 [アクセス2016-12-03] 。
- ↑ A.J. マーティン 、 骨のない恐竜:恐竜の生活微量化石によって明らかにされた 、ロンドン:Pegasus Books、2014、s。 66–67、ISBN 978-1605984995 。
- ↑ A. ロミリオ 、 恐竜のスタンピードはそのトラックで停止しました 、「オーストラリア科学」、2、36、2015、s。 24–27 、ISSN 1442-679x 。
- ↑ 広告。 Shapiro 、 決してなかった偉大な恐竜のスタンピード? 、 2013 [アクセス2018-05-07] 。
- ↑ マレク・ジェディニアック: 先史時代の動物 – 恐竜 。 www.kzp.pl. [アクセス2019-11-28]。 ( pol。 )) 。
- ↑ ガリミムス 。 colnect.com。 [アクセス2019-11-28]。 ( pol。 )) 。

Recent Comments