ババコティア – ウィキペディア、無料百科事典
ババキティア – マダガスカルのレミカルグループ(Strepsirrhini)からの絶滅したタイプの培地サイズの哺乳類。単一の種をカバーします: ババコティアラドフィライ 。一緒に Palaeopropithecus 、 Archaeoindris 私 Mesopropithecus 英語で呼ばれる古生理学科を作成します ナマケモノのキツネザル ( 怠惰 怠zyを意味します)。ジャンル名 ババキティア マルガスカのインドリスの名前から来ています、 ババコト 、近い相対的な古生物生物科。怠zyでより高度な偉大な代表者を連想させる遅い小さな雌丸の木の間の移行段階のように見える形態学的特徴のため、属は両方のグループ間の関係を決定するのを助けました。
ババコティアラドフィライ そして、他のすべてのPalaeopropithecidaeは、現代の怠zyな人々に見られる多くの特徴を分けます。これは並行進化の例です。彼らは、強い前腕、湾曲した指、腰、重要な可動特性を持つジャンプジョイントを持っています。頭蓋骨は、インディリダ科よりも強い構造を示していますが、偉大な古生物岩科ほどではありません。歯は、怠zyまたはインディリダエと比較して、他のキツネザルで見られるものに似ています。動物はマダガスカルの北部に住んでおり、少なくとも2つの他の古生物術科種とその発生範囲を共有しています。 Palaeopropithecus巨大 私 Mesopropithecus dolichobrachion 。 ババコティアラドフィライ 彼は主に葉を消費しましたが、果物や硬い種を食べることもできました。彼は彼らのサブフォシルの遺物でのみ知られており、島に男性が到着した直後に死ぬ可能性があります。ただし、十分な放射性炭素年代測定が実行されなかったため、確実性はありません。
ジャンル名 ババキティア 彼はマルガス語から来ています。近親者であるインドリスの通常の名前からソースを取得します ババキティア 、この言語で呼ばれます ババコト 。次に、種のepithet ラドフィル 彼は洞窟地図を作ったグロトワズのジャン・ラドフィラオのフランスの数学を称え、属の最初の遺跡が見つかった [3] 。
ババコティアラドフィライ 彼は属の唯一のメンバーです ババキティア 。それは古生物の科に属し、怠zyを連想させる他の3つのタイプのレモンもカバーしています。 Palaeopropithecus 、 Archaeoindris 私 Mesopropithecus 。この家族は、すべてのマダガスカルのキツネザルを含むレムリフォルメスの下位に配置されています [4] [5] 。
最初のサブフォシールが残ります ババコティアラドフィライ 1981年と1986年から1986年にかけてのジャンラドフィラオと2つのアングロマルガス偵察遠征の発見の後に発生した一連の遠征中に発見されました。 [6] [7] 。発見の第二の波は、生物人類学者のエルウィン・サイモンズによって80年代に始まりました [8] 、1988年にマダガスカルの北にあるアンカラン・マシフのアント・シランドーハの洞窟を襲いました。 [2] 。 Palaeopropithecidaeの代表として迅速に識別されました [3] 、 ババキティア 一緒に Mesopropithecus 彼女は、古植物類の科、古科、そして現在住んでいる屋内の間の親族の議論の始まりに貢献しました。 Archaeolidaeには、インディダエで遭遇したものを非常に連想させる頭蓋骨がありました。一方、より大きな古生物術科は、現在のインディリダエに似た歯を保持していますが、より固体で専門的でした。 ババキティア 私 Mesopropithecus 彼らはインディリダエと同じ歯を持っていただけでなく、同様の頭蓋骨も持っていました。これは、レニウカに似たキツネザルが今日のインディリダエ科に最も近く、アルケリミダエが最も近い外部グループであることを証明しています [9] 。さらに、発見 ババキティア 元のインディリダエは、現代の代表者のように、垂直の登山家が吊り下げ姿勢で餌をやり、おそらくジャンパーでのみ垂直の登山家が木から木へと動くジャンパーではないことを示すのに役立ちました。 [十] 。
ババキティア そして、filogenezylemurów [11] [12番目] [13]
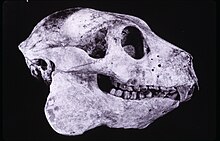
重量は16〜20 kg、 ババコティアラドフィライ 彼女は中型のキツネザルに属していました。偉大な古生物生物科よりもはるかに小さかった Archaeoindris どうにか Palaeopropithecus しかし、彼女は小さな古生物生物科のサイズを超えました。 Mesopropithecus [4] [5] または彼女と関連するタイプ デュゲントニア 。さまざまな種 Archaeolemur より小さく達成された( A.メジャー )より大きくなる( A.エドワードティ ) 体重。 Jungersと同僚は、それが16.2 kgの重さを量ったと言います – 今日のパビアンの男性と同じくらい [14] 。
多くの機能に関しては、SO -Calledへの適応の間接レベルに達しました大頭と小さなキツネザルの間の挙動の懸濁液は古生物生物科 [5] [2] [13] [9] [15] 。それらには、腰または足首の関節に大きな可動性が含まれ、脊椎、骨盤、手足の構造を含む他の適応が含まれます [15] 。前足は後部より20%長く、これは約119の上肢と下肢(膜間指数)の長さの比率が高い、u Mesopropithecus それは97から113の間でした。これは、オーバーラップが怠zyとの収束を示唆しています [15] [16] 。肩と構造の指標(平均最大の最大寸法と平均最大大腿骨寸法に100を掛けた比率を表す)は115です。 クリエイティブ 、そして今日のサルの中で、この指標はゴリラで118です。肩の骨の長さは質量に修正されたのは70.8 mm*kgです -0.5 (最も近い結果は次のとおりです Mesopropithecus doichbrachion )。大腿骨の骨の場合、この値は61.7 mm*kgです -0.5 、代表者でも同様の結果が発生します Palaeopropithecus 私 Mesopropithecus [14] 。散歩は減りました。指は細長く湾曲し、食物摂取に適応しました。言及されたサスペンサーの動作を提案しました [15] [17] 。近位指紋の平均曲率は59.1°C(灰色のウールのような)でした。足、手のように、比較的長い [14] しかし、他の古生物生物科と同様に、登山と吊り下げに適応して、彼らは減少しましたが、ジャンプではありません(室内のような) [4] [5] [2] 。 1999年に見つかった手首の骨は、種が垂直の位置に登ったことを示しました [18] 。さらに、セミクラウシングチャネル分析 [19] 、腰椎とその辛い付属物は、ゆっくりした動きと今日の素晴らしい猿のように登るための適応を示しています( アントプロノグレード 、それは4つの手足で垂直サポートをつかむことです)が、必ずしも怠zyな吊り下げ、垂直接着、またはジャンプではありません [20] 。 ババキティア ゆっくりとした登山家に似ていました ロリス 、怠zyのように動作サスペンサーのいくつかの機能を示しています [8] 。
すべてのPalaeopropithecidaeが特徴付けられています [5] 。しかし、偉大なキツネザルにも見られる機能にもかかわらず [9] 頭蓋骨 ババキティア それはまだインディリダエを連想させます [4] 。他のLemuri Palaeopropithecidaeと共有される頭蓋骨の特徴には、比較的小さなアイソケット、強力な接合弓、およびほとんど長方形の硬口蓋が含まれます [5] 。視覚チャネルの相対的なサイズと比較して小さなアイソケットは、キツネザルの典型的な低い視力を示唆しています [21] 。頭蓋骨の長さは平均144 mmです [2] 。アイソケットの直径と体重と頭蓋骨の長さの比較(オスゴマティック後、さまざまな霊長類のこれらの値は、まっすぐな線についての座標系に配置されています) ババキティア 夜の種とはほど遠いですが、毎日のサルとも異なります [14] 。
この種の歯のパターンは、他の古生物生物科およびインドリダエと違いはありませんでした:顎で2.1.2.3、顎で1.1.2.3 [5] [9] 2.1.2.3 I 2.0.2.3 [2] 。永久歯の歯の1つが実際にはガラスなのかチョッパーなのかは説明されていません。 [22] 。いずれにせよ、牙やチョッパーの不足は、6つのsotarのより典型的なものではなく、歯の紋章と呼ばれる4倍の前歯のグループを引き起こします [2] 。 4歯の櫛も実行されました Mesopropithecus [14] 。 ババコティアラドフィライ それはインディリダエとわずかに異なり、わずかに長く伸びた施設を持っています。頬の歯には、広い切断端と鋸歯状のエナメル質がありました [2] 。歯電子顕微鏡 B.ラドフィル そして素晴らしい クリエイティブ どちらの場合も、彼女は毛皮をとませることを証言する溝を明らかにしました。これは追加の歯の櫛機能であるように思われますが、それが自分の毛皮のトイレであろうと他の個人のトイレに関するものであろうと、現在の知識の状態で知ることはできません。一方で Babot 彼女は足のもう一方の指に毛皮をとかすために爪を保存しました [14] 。
地理的分布と生態学 [ 編集 | コードを編集します ]
他のすべてのキツネザルのように、 ババコティアラドフィライ それはマダガスカルのエンデムに属します。彼女の遺体は、レザーブ・スペシアレ・ダンカナのアンカラン・マッシフのカルスト洞窟と、北と北西の島の北と北西部を覆っていることを示しているアンジョヴェにのみ発見されました。 [5] [2] [23] 。特にマダガスカルの大部分が森林で覆われていた時期に、このテーラー – サッデンの限られた範囲は、生息地の詳細、ガウスの原則、またはその他の未知の要因に起因する可能性があります。 [24] 。彼らは彼女に同情しました Palaeopropithecus最大 私 Mesopropithecus dolichobrachion [8] [25] [26] 。
歯の体の大きさ、モル形態、顕微鏡分析に基づいて、それは認識されています ババコティアラドフィライ 緑豊かな食事のために、その食事を果物と硬い種で補完する [4] [13] [21] [27] (どうやって Palaeopropithecus )、時々それは種や他の硬い食べ物を食べると考えられています [28] 。動物の歯の微小腫瘍の研究は、葉や種子と矛盾していませんが、同様のメニューを持つ他のキツネザルと比較して、それらにはわずかな違いがあります。これは小さな分離を示しています [14] 。しかし、ゴッドフリーと同僚の研究は、帰属の可能性を評価しました ババキティア 0.02の草原の場合、および果物は0.98で発生します。また、彼女は彼女の食事を種子で豊富に補充したことが認識されていました(おそらく、彼女は古生物岩科で最も多くの種子を食べたかもしれません)。今日、同様の食事は、ゴリラ西部、ボルネアオランウータン、サキグレイ、サキビアウォーリア、および Cacajao Rubicunda 。歯の消費の厚い痕跡 ババキティア そして、それほどではないが、彼女の家族の他の代表者は、爪の歯を飾るものに似ています。 Palaeopropithecidaeも多くの葉を消費しました [28] 。したがって、すべてのPalaeopropithecidaeで、したがって ババコティアラドフィライ 、胸から止まった後、最初の乾季に若者の生存を増加させたインドリダエ科のように、永久歯が早期に噴火しました。 [15] 。
動物は地質学的に比較的最近死んだので [23] 。 ババコティアラドフィライ 彼女は完新世の間に住んでいた。おそらく私たちの時代の第2千年紀の間に、人々の島に到着した直後に彼女は姿を消したと考えられています [5] 。ただし、このケースで使用されている唯一の放射性炭素年代測定は、紀元前3100年から2880年の発見の年齢をマークしました。 [29] 。
- ↑ M.C.マッケナ、S.K。ベル: 哺乳類の分類:種レベルを超える 。コロンビア大学出版局、1997年、s。 335. ISBN 0-231-11013-8 。 ( 。 )) 。
- ↑ a b c d そうです f g h 私 第7章:四角い化石雌牛。 W:L.R。ゴッドフリー、W.L。ジュンナー: 霊長類の化石記録 。ケンブリッジ大学出版局、2002年、s。 97 –121。 ISBN 978-0-521-66315-1 。 ( 。 )) 。
- ↑ a b L.R.ゴッドフリー、E.L。サイモンズ、p。チャタス、B。ラコトサミマナナ。 新しい化石のキツネザル( ババキティア 、霊長類)北部マダガスカルから 。 「科学アカデミーの報告」。 81、s。 81–87、1990。 ( 。 )) 。
- ↑ a b c d そうです 第3章:絶滅のキツネザル。 W:R.A。 Mittermeier、W.R。Konstant、F。Hawkins、E.E。Louis、O。Langrand: マダガスカルのキツネザル 。 S.D.ナッシュ。 WYD。 2番目。保護国際、2006年、s。 37–51。 ISBN 1-881173-88-7 。 ( 。 )) 。
- ↑ a b c d そうです f g h 私 Palaeopropithecidae科:ナマケモノのキツネザル。 W:R.M。 Nowak: ウォーカーの世界の哺乳類 。 WYD。 6.ジョンズ・ホプキンス大学出版局、1999年、s。 89 –91。 ISBN 0-8018-5789-9 。 ( 。 )) 。
- ↑ J.M.ウィルソン、P.D。スチュワート、G.-S。ラマンガソン、AMデンニング私がイニニ。 アンカラナ、N。マダガスカルのcrown冠されたキツネザルの生態と保存サンフォードのキツネザル、その他の同所性、および亜化石雌師に関するメモ 。 「folia primatologica」。 52、s。 1–26、1989。doi: 10.1159/000156379 。 ( 。 )) 。
- ↑ J …ウェッソン、。神は呼ばれています、それ。 Seeity、P。サルトリーもネニーニです。 Ankarana、N。Madagascarの過去と現在のLemur Fauna 。 「霊長類の保全」。 16、s。 47–52、1995。 ( 。 )) 。
- ↑ a b c L.R.ゴッドフリー、W.L。ジョンガー。 マダガスカルの絶滅したナマケモノ 。 「進化人類学」。 12(6)、s。 252–263、2003。doi: 10.1002/evan.10123 。 ( 。 )) 。
- ↑ a b c d 第4章:絶滅のキツネザル。 W:R。A. Mittermeier、I。Tattersall、W.R。Konstant、D.M。マイヤーズ、R.B。マスト: マダガスカルのキツネザル 。 S.D.ナッシュ。 WYD。 1番目。 Conservation International、1994、s。 33–48。 ISBN 1-881173-08-9 。 ( 。 )) 。
- ↑ W.L. Junners、L.R。ゴッドフリー、E.L。サイモンズ、P.S。 chatrath i inni。 の系統および機能的親和性 ババキティア (霊長類)、マダガスカル北部の化石雌牛 。 「国立科学アカデミーの議事録」。 88(20)、s。 9082–9086、1991。doi: 10.1073/pnas.88.20.9082 。 bibcode: 1991pnas … 88.9082J 。 ( 。 )) 。
- ↑ J.E. Horvath、D.W。ワイスロック、S.L。 Embry、I。FiorentinoI inni。 系統発生ツールキットの開発と応用:マダガスカルのキツネザルの進化の歴史の解決 。 「ゲノム研究」。 18(3)、s。 489–499、2008。doi: 10.1101/gr.7265208 。 PMID: 18245770 。 [アクセス2010-02-24]。 [アーカイブ 住所 ]。 ( 。 )) 。
- ↑ L.オーランド、S。カルヴィニャック、C。シュネーベン、C.J。 Doudy I inni。 絶滅の巨大なキツネザルからのDNAは、アルケオレムリドを現存するインドリッドにリンクしています 。 「BMC進化生物学」。 8、s。 121、2008。doi: 10.1186/1471-2148-8-121 。 PMID: 18442367 。 ( 。 )) 。
- ↑ a b c L.R.ゴッドフリー、W.L。ジュンナー: マダガスカルの自然史 。シカゴ大学出版局、2003年、s。 1247–1252。 ISBN 0-26-30306-3 。 ( 。 )) 。
- ↑ a b c d そうです f g ウィリアム・L・ジンジャー、ローレ・R・ゴッドフリー、エルウィン・L・サイモンズ、ロシュナ・E・ワンダーリッチ、ブライアン・G・リッチモンド、プリティジット・S・チャットラス:マダガスカルのガント・絶滅・キツネザルの原体形成と行動。 W:J。マイケルプラヴカン、リチャードF.ケイ、ウィリアムL.ジャンガー、カレルP.ヴァンシャイク: 霊長類の化石記録における行動の再構築 。 2002年、s。 371-411、SERIA:霊長類の進歩。 ISBN 978-1-4615-1343-8 。 [アクセス2013-11-30]。
- ↑ a b c d そうです 第6章:キツネザル:古いものと新しい。 W:E.L。サイモンズ: マダガスカルにおける自然な変化と人間の影響 。 Smithsonian Institution Press、1997、s。 142–166。 ISBN 978-1-56098-682-9 。 ( 。 )) 。
- ↑ 第8章:サブファーティールキツネザル。 W:L.R。ゴッドフリー、W.L。ジャンカー、K.E。リード、E.L。サイモンズ、P.S。 Chatrath: マダガスカルにおける自然な変化と人間の影響 。 Smithsonian Institution Press、1997、s。 218–256。 ISBN 978-1-56098-682-9 。 ( 。 )) 。
- ↑ W.L. Junners、L.R。ゴッドフリー、E.L。サイモンズ、P.S。 chatrath。 絶滅したナマケモノのキツネザル(霊長類、古植物科)における眼球の曲率と位置的行動 。 「国立科学アカデミーの議事録」。 94(22)、s。 11998–12001、1997。doi: 10.1073/pnas.94.22.11998 。 PMID: 11038588 。 bibcode: 1997pnas … 9411998J 。 ( 。 )) 。
- ↑ M.W.ハムリック、E.L。サイモンズ、W.L。ジョンガー。 マラガシーガント亜鏡皮の新しい手首の骨 。 「Journal of Human Evolution」。 38(5)、s。 635–650、2000。doi: 10.1006/jhev.1999.0372 。 PMID: 10799257 。 ( 。 )) 。
- ↑ A.ウォーカー、T.M。ライアン、M.T。シルコックス、E.L。サイモンズIインニ。 半円形の運河システムと運動:絶滅の葉巻とロリソイドの場合 。 「進化人類学」。 17(3)、s。 135–145、2008。doi: 10.1002/evan.20165 。 ( 。 )) 。
- ↑ L.J.シャピロ、C.V.M。失敗、L.R。ゴッドフリー、W.L。 Jungers I Inni。 絶滅したマラガシーストレプシルハインにおける腰椎椎骨の形態計測分析 。 「American Journal of Physical Anthropology」。 128(4)、s。 823–839、2005。Doi: 10.1002/JPA 。 PMID: 16110476 。 ( 。 )) 。
- ↑ a b 第3章:マダガスカルのサブ化石雌ツの生態学とext骨。 W:L.R。ゴッドフリー、W.L。 Jungers、G.T。シュワルツ: Lemurs:生態学と適応 。 Springer、2006、s。 41 –64。 ISBN 978-0-387-34585-7-7 。 ( 。 )) 。
- ↑ 第7章:歯。 W:F。アンケルシモンズ: 霊長類の解剖学 。 WYD。 3. Academic Press、2007、s。 224–283。 ISBN 0-12-372576-3 。 ( 。 )) 。
- ↑ a b 第4章:夜行性レムムリフォルム。 W:R.W。Sussman: 霊長類の生態学と社会構造 。 Pearson Custom Publishing、2003、s。 107–148。 ISBN 978-0-536-74363-3 。 ( 。 )) 。
- ↑ D.A.バーニー、H.F。ジェームズ、F.V。 Grady、J。RafamantanantsoaI Inni。 環境の変化、絶滅、人間の活動:NWマダガスカルの洞窟からの証拠 。 「生物地理学のジャーナル」。 24(6)、s。 755–767、1997。doi: 10.1046/j.1365-2699.1997.00146.x 。 ( 。 )) 。
- ↑ l。 Godphyy、Jane San。シーミアン人、ポール。 アンカラナ:マダガスカルの過去の窓 。 「レマーニュース」。 2、s。 16–17、1996。 ( 。 )) 。
- ↑ JainTin。WeAreson、Lin。ゴッドファシー、それ。シーミアン人、ポール。 Ankarana、N。Madagascarの過去と現在のLemur Fauna 。 「霊長類の保全」。 16、s。 47–52、1995。 ( 。 )) 。
- ↑ K.L. Raffyy、M.F。 Teafford、W.L。ジョンガー。 化石皮のモルマイクロヤー:食事の推論の解決を改善する 。 「Journal of Human Evolution」。 43(5)、s。 645–657、2002。doi: 10.1006/jhev.2002.0592 。 PMID: 12457853 。 ( 。 )) 。
- ↑ a b Laurie R. Godfrey、Gina M. Semprogon、William L. Junners、Michael R. Sutherland、Elwhen L. Simouns&Nichos Solonias。 絶滅したキツネザルでの歯の使用摩耗:食事とニッチの違いの証拠+ 。 「Journal of Human Evolution」。 47、s。 145-169、2004。科学直接。エルゼビア。 ( 。 )) 。
- ↑ E.L.サイモンズ、D.A。バーニー、P.S。 Chatrath、L.R。 godfrey i inni。 AMS 14 cマダガスカル北部のアンカラナ・マッシフの洞窟からの絶滅のキツネザルの日付 。 「第四紀研究」。 43(2)、s。 249–254、1995。Doi: 10.1006/QRES.1995.1025 。 bibcode: 1995キーベス..43..249S 。 ( 。 )) 。
Recent Comments