トランジショナルフォーム-Wikipedia、無料百科事典

トランジショナル(間接)フォーム – 2つの体系的なグループの特徴をつなぐ生物の形態。
チャールズ・ダーウィンは、以前の地質時代に間接的な形の発生を想定していました 種の形成について [初め] 。自然選択の結果としての進化理論の創造者は次のように書いています。
最終的に、私の理論が正しければ、1つの期間ではなく過去全体を考慮に入れて、特定のグループのすべての種を可能な限り密接に組み合わせることができるように、間違いなく無数の間接的な品種が存在する必要がありますが、自然選択プロセス自体は、よく言及しているように、親の形と中間細胞の破壊に常に努力しています。
種の形成に関するカロル・ダーウィンは、 Szymon DicksteinとJózefNusbaum、州農業およびLeśne出版社、ワルシャワ1959、p。173。
ダーウィンの作品が作成されましたが、絶滅した生物の化石はほとんどありませんでした。しかし、それは出版物の2年後に発見されました Archeopteryx 、体はしばしば最初の既知の鳥と呼ばれ、2つの大きな種の特徴を示しています。翌年、化石が発見され、今日の多くの種のグループ間で間接的な特徴を持つ生物の存在を確認しました [初め] 。
よくある間違いは、進化生物学が生物の進化史に「ミッシングリンク」を探しているという信念です。古生物学者は、進化の変化に起因する枝を考慮して、主に系統樹を構築するために化石記録を使用しています [2] 。 「ミッシングリンク」の概念は非常に示唆的であり、生物の主要なグループの始まりをキャプチャできるようにする以前は無関係でまれな移行フォームに関連しています [3] 。概念の出現と普及は、進化の理論よりも古く、存在の大きな連鎖の概念を指します。これは、物質と形而上学の世界の要素を存在の連鎖と呼ばれる階層構造に編成します。 18世紀から19世紀にかけて、エンティティの連鎖の概念は生物学の進化的思考に関連しており、その反りにつながりました [2] 。進化の進化の知覚は、進歩の仮定でより低い形態からより高い形に変化することは、一連の枝と多様性の増加としてのみ説明できる共通起源の考えを否定します [4] 。 「ミッシングリンク」の重要な部分は長い間発見され、このステータスを失いました。
移行形態と進化の理論 [ 編集 | コードを編集します ]
| 1850 |  |
| 1900 |  |
| 1950年 |  |
| 2002年 | 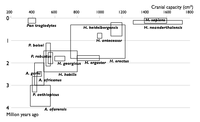 |
| 科学を示すチャート – 選択された年に有名な人類の種。各種は、特定の種の発見の頭蓋腔の体積と発見の限界日を示す長方形として示されています。図シーケンスは、新しい発見を通じて、以前知られている種間の空間の充填を示しています。 | |
19世紀には、進化に関する見解は、ダーウィニズム、ニアウィニズム、ラマキズム、ネオマルーキズムなど、偉大な理論の一部として形作られました。 H. de Vriea、C。Corens、およびE. Tschermak Mendelの1900年のE. Tschermak Mendelの権利(研究者のそれぞれが独立してこれを行った)による繰り返し発見は、遺伝のメカニズム、遺伝学の発展、進化の新しい見解を理解することにつながりました。 1900年から1930年の年は、E。DodsonAsによって定義されました 進化的不可知論の時代 。 E. Mayrのみが、自然選択の理論を備えた人口遺伝学、生態学、生物地理学の一部として獲得した知識をまとめ、地理的専門の理論を策定しました。これは、微小進化を説明する完全な合成です。古生物学的証拠は彼女にとって重要ではありませんでした [5] 。古生物学的データはによって使用されました
G.シンプソン、本の中で 進化のテンポとモード 1944年に出版されました。古生物学者の中で、ダーウィンの理論はほとんど支持者を獲得しませんでした。ネオラマリズムの見解は支配的であり、大きな体系的なグループが作成されたと仮定していました。古生物学者であるシンプソンは、マクロ進化の分野におけるダーウィン理論を伴う遺伝学と古生物学の分野で統合されました [6] 。量子進化の決定を使用して、マルチgeneen相続モデルを使用して、シンプソンは、適応の1つのトップから次のトップへの大きな集団の移行を説明しました。連続したバランス状態間の迅速な移行は、集団の十分に高い変動性で可能であり、環境の変化と身体内の変化(例:突然変異)の両方の結果として発生する可能性があります。いくつかの標準偏差の表現型の変動性 [7] 。このようにして、なぜ種の大規模なグループ間の遷移形成が化石の形で動作しない理由が説明されました [5] 。
現在、進化の理論は、地質学的時間と移行形態があるべき場所を非常に正確に決定することを可能にします。これらの予測は、古生物学の検索練習で大規模に使用されます。このように発見された化石の1つはです 刺繍 、マンディック魚と初期の四足動物の間の移行型 [8] 。 20世紀の初めには、水からrawい出るひれで動物の存在のアイデアが登場しました。 1930年代には、指名された動物の化石がグリーンランドで発見されました Ichthyostega 、後期のデウォンから。魚と4脚の動物との間の進化の過程に関連する多数の仮説と憶測が文献に広がっています。 21世紀初頭からの発見がこれらの仮説をテストすることを許可されたことだけでした。 [9] 。
- ↑ a b シドパーキンス。 stephyphen; ‘s by-hyphen;’ s step進化:ギャップのマイニング:過渡期の化石は見つけるのが最も難しいですが、時には最高の物語を語ることもあります 。 「科学ニュース」。 175(3)、s。 30–33、2009。Doi: 10.1002 / scin.2009.5591750320 。 ISSN 00368423 。 ( 。 )) 。
- ↑ a b ルイーズ・S・ミード。 過渡的な形についての私たちの思考を変えます 。 「進化:教育とアウトリーチ」。 2(2)、s。 310–314、2009。doi: 10.1007/s12052-009-0126-3 。 ISSN 1936-6426 。 ( 。 )) 。
- ↑ エリック・アールバーグ、ジェニファー・A・クラック 古生物学:水から陸への確固たるステップ 。 “自然”。 440(7085)、s。 747–749、2006。doi: 10.1038/440747a 。 ISSN 0028-0836 。 ( 。 )) 。
- ↑ エマニュエル・リガート、アレッサンドロ・ミネリ。 存在の偉大なチェーンはまだここにあります 。 「進化:教育とアウトリーチ」。 6(1)、s。 18、2013。doi: 10.1186/1936-6434-6-18 。 ISSN 1936-6434 。 ( 。 )) 。
- ↑ a b Urbanek Jerzy。 20世紀の生物学 – 主な流れ 。 「コスモス」。 49(3(248))、s。 305–319、200。
- ↑ エルンスト・メイール、ウィリアム・B・プロヴァイン: 進化的統合:生物学の統一に関する視点 。ハーバード大学出版局、1998年、s。 153–173。 ISBN 978-0-674-27226-2 。 ( 。 )) 。
- ↑ マーク・カークパトリック。 継続的な遺伝文字における量子進化と句読点平衡 。 「アメリカの自然主義者」。 119(6)、s。 833–848、1982。
- ↑ エドワード・B・デシュラー、ニール・H・シュビン、ファリッシュ・A・ジェンキンス。 デボン紀のテトラポッドのような魚とテトラポッドボディプランの進化 。 “自然”。 440(7085)、s。 757–763、2006。doi: 10.1038/nature04639 。 ISSN 0028-0836 。 ( 。 )) 。
- ↑ ジェニファー・A・クラック。 魚と星の遷移:新しい化石と解釈 。 「進化:教育とアウトリーチ」。 2(2)、s。 213–223、2009。doi: 10.1007/s12052-009-0119-2 。 ISSN 1936-6426 。 ( 。 )) 。


Recent Comments