テロメラーゼRNA要素 – Wikipedia
テロメラーゼRNA要素(テロメラーゼRNAようそ、英: telomerase RNA component、略称: TERC)は真核生物に存在するノンコーディングRNAで、テロメアの伸長に用いられる酵素 テロメラーゼの構成要素である[2][3]。テロメラーゼRNA構成要素、テロメラーゼRNA成分などとも呼ばれる。TERCは、テロメラーゼによるテロメアの複製(逆転写)の際の鋳型として機能する。TERCの配列と構造は脊椎動物、繊毛虫類、酵母の間で大きく異なるが、鋳型配列に近接した5’末端側にシュードノット構造が存在することは共通している。脊椎動物のTERCの3’末端にはH/ACA型snoRNA様ドメインが存在している[4][5][6]。
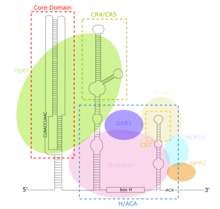
TERCは長鎖ノンコーディングRNA(lncRNA)の1つであり、その長さは繊毛虫類では約150ヌクレオチド、脊椎動物では400–600ヌクレオチド、酵母では約1300ヌクレオチドである[7]。成熟したヒトTERC(hTR)は451ヌクレオチドである[8]。TERCは広範囲の二次構造を持ち、4つの主要な保存されたドメインからなる[7]。TERCの5’末端に位置する最大のドメインであるコアドメインには、テロメアの鋳型となるCUAAC配列が含まれている。その二次構造は、鋳型配列を含む大きなループ、ループを閉じるP1ヘリックス、P2/P3シュードノットからなる[9]。コアドメインと保存されたCR4/CR5ドメインがTERTと結合し、テロメラーゼのin vitroでの触媒活性にはこれらのドメインのみが必要である[10]。TERCの3’末端には保存されたH/ACAドメインが存在し、一本鎖のヒンジで連結された2つのヘアピン構造と3’末端の一本鎖ACA配列からなる[8]。H/ACAドメインはジスケリン、GAR1、NOP10、NHP2と結合し、H/ACAリボヌクレオタンパク質複合体を形成する[9]。TERCの3’末端には保存されたCR7ドメインも存在し、TCAB1が結合する3ヌクレオチドのCABボックス(カハール体への局在を担う)を含んでいる[9]。

テロメラーゼはリボヌクレオタンパク質からなるポリメラーゼで、テロメアの末端にTTAGGGのリピート配列を付加することで末端を維持する。リピート配列は、真核生物の種によって差異が存在する。テロメラーゼには逆転写酵素活性を持つタンパク質要素(TERT)と、テロメアリピートの鋳型として機能するRNA要素(TERC)が含まれている。脊椎動物型TERC配列の50位付近に存在するCCCUAA配列が鋳型として機能する。テロメラーゼの発現は細胞老化と関係している。出生後の体細胞では通常抑制されており、テロメアは短縮してゆく。体細胞でのテロメラーゼの発現調節の異常は発がんに関与している可能性がある。マウスでの研究からは、テロメアリピートの新規合成が二本鎖切断部位でも生じている可能性が示されており、テロメラーゼが染色体修復にも関与している可能性が示唆されている[11]。TERCのホモログはトリヘルペスウイルスにも存在する[12]。
TERCのコアドメインは、TERTがTTAGGGからなるテロメアリピートを合成する際のRNA鋳型を含んでいる[9]。他のリボヌクレオタンパク質と異なり、テロメラーゼではタンパク質のTERTが触媒活性を持つ一方、lncRNAであるTERCは構造的な役割を持ち、リボザイムではない[13]。In vitroでテロメラーゼの触媒活性を再構成するには、TERCのコア領域とTERTで十分である[9][10]。TERCのH/ACAドメインはTERCを安定化するジスケリン複合体(DKC1、GAR1、NOP10、NHP2)をリクルートし、テロメラーゼ複合体の形成と全体的な触媒活性を向上させる[9]。CR7ドメインはテロメラーゼをカハール体へ局在させるTCAB1と結合し、テロメラーゼの触媒活性をさらに向上させる[9]。テロメラーゼ活性やTERTの発現がみられない細胞においても、TERCは普遍的に発現している[14]。そのため、TERCのTERT非依存的なさまざまな役割が提唱されている。TERC結合モチーフを含む14の遺伝子はTERCによる直接的な転写調節を受けており、RNA-DNA三重鎖形成によって発現が増加する。TERCによるLIN37、TRPG1L、TYROBP、USP16のアップレギュレーションはNF-κB経路を刺激し、炎症性サイトカインの発現と分泌を増加させる[15]。
スプライソソームによってイントロンから組み立てられる大部分のlncRNAとは異なり、ヒトTERC(hTR)は、ゲノム上3q26.2の遺伝子座[16]に位置する専用のプロモーターからRNAポリメラーゼIIによって直接転写される[8]。成熟したhTRは451ヌクレオチドの長さであるが、細胞内の定常状態のhTR転写産物の約1/3にはゲノムにコードされた10ヌクレオチド程度の3’テールが存在する。こうした伸長型hTRの大部分には、さらにオリゴ(A)からなる3’末端の伸長が存在する[8]。3’テールを持つ未成熟型hTRから451ヌクレオチドの成熟型hTRへのプロセシングは、3′-5’エキソリボヌクレアーゼによる直接的な分解、もしくはPAPD5によるオリゴアデニル化、3′-5’エキソリボヌクレアーゼPARNによる3’オリゴ(A)テールの除去、そして3′-5’エキソリボヌクレアーゼによる分解、という間接的な経路によって行われる[8]。伸長型のhTRはエキソソームによっても分解される[8]。
hTR転写産物の5’末端もプロセシングを受ける。TGS1による5’キャップの2,2,7-トリメチルグアノシンへのメチル化は、hTRの成熟を阻害する[17]。hTR転写時のH/ACAドメインへのジスケリン複合体の結合は、転写終結を促進する[8]。こうしたhTRの成熟の活性化または阻害を行うさまざまな競合的経路の相対的速度の制御は、テロメラーゼ活性の全体的な調節の重要な要素となっている。
臨床的意義[編集]
TERCの機能喪失型変異は、さまざまな変性疾患と関係している。TERCの変異は、先天性角化異常症[18]、特発性肺線維症[19]、再生不良性貧血、骨髄異形成症候群と関係している[9]。TERCの過剰発現と不適切な調節は、さまざまながんと関係している。hTRのアップレギュレーションはHPVの感染による前がん状態の患者で広く観察される[20]。TERCの過剰発現はマルディウイルスの発がん性を高める[21]。TERCの過剰発現は胃がんで観察される[22]。TERCの過剰発現は2型糖尿病や多発性硬化症などの炎症性疾患でも観察され、TERCによってNF-κB炎症経路が活性化される[15]。
TERCは骨粗鬆症に対する保護効果が示唆されており、TERCの発現の低下は骨形成を低下させる[23]。TERCはさまざまながんで過剰発現しているため、がんのバイオマーカーとしての利用可能性が研究されている。肺扁平上皮がんにおいては、有効なバイオマーカーであることが示されている[24]。

Recent Comments