Thioredoxinreduktase – Wikipedia
Thioredoxinreduktasen ((TR, TrxR) (EG 1.8.1.9) sind die einzigen bekannten Enzyme, die Thioredoxin (Trx) reduzieren.[1] Es wurden zwei Klassen von Thioredoxinreduktase identifiziert: eine Klasse bei Bakterien und einigen Eukaryoten und eine bei Tieren. In Bakterien katalysiert TrxR auch die Reduktion von Glutaredoxin-ähnlichen Proteinen, die als NrdH bekannt sind.[2][3][4] Beide Klassen sind Flavoproteine, die als Homodimere fungieren. Jedes Monomer enthält eine FAD-Prothesengruppe, eine NADPH-Bindungsdomäne und ein aktives Zentrum, das eine redoxaktive Disulfidbindung enthält.[5]
Zelluläre Rolle[edit]
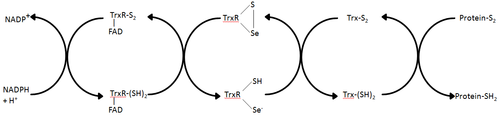
Thioredoxinreduktase ist das einzige bekannte Enzym, das die Reduktion von Thioredoxin katalysiert[1] und ist daher eine zentrale Komponente im Thioredoxinsystem. Zusammen mit Thioredoxin (Trx) und NADPH beschreibt dieses System am allgemeinsten die Bildung reduzierter Disulfidbindungen in Zellen. Elektronen werden über TrxR aus NADPH entnommen und auf das aktive Zentrum von Trx übertragen, das anschließend Proteindisulfide oder andere Substrate reduziert.[6] Das Trx-System existiert in allen lebenden Zellen und hat eine Evolutionsgeschichte, die an DNA als genetisches Material gebunden ist, Abwehr von oxidativen Schäden durch Sauerstoffmetabolismus und Redoxsignalisierung mit Molekülen wie Wasserstoffperoxid und Stickoxid.[7][8]
Vielfalt[edit]
Zwei Klassen von Thioredoxinreduktase haben sich unabhängig voneinander entwickelt:
Diese beiden Klassen von TrxR haben nur ~ 20% Sequenzidentität in dem Abschnitt der Primärsequenz, in dem sie zuverlässig ausgerichtet werden können.[5] Die Nettoreaktion beider TrxR-Klassen ist identisch, aber der Wirkungsmechanismus ist jeweils unterschiedlich.[9]
Menschen exprimieren drei Thioredoxinreduktase-Isozyme: Thioredoxinreduktase 1 (TrxR1, cytosolisch), Thioredoxinreduktase 2 (TrxR2, mitochondrial), Thioredoxinreduktase 3 (TrxR3, testis-spezifisch).[10] Jedes Isozym wird von einem separaten Gen kodiert:
Struktur[edit]
E coli[edit]
Im E coli ThxR gibt es zwei Bindungsdomänen, eine für FAD und eine für NADPH. Die Verbindung zwischen diesen beiden Domänen ist ein zweisträngiges antiparalleles β-Faltblatt.[11] Jede Domäne ist den analogen Domänen in Glutathionreduktase und Lipoamiddehydrogenase sehr ähnlich, aber ihre relative Orientierung dieser Domänen in ThxR ist um 66 Grad gedreht.[11] Dies wird im nachstehend beschriebenen Enzymwirkungsmechanismus von Bedeutung. ThxR homodimerisiert mit der Grenzfläche zwischen den beiden Monomeren, die aus drei Alpha-Helices und zwei Schleifen bestehen.[11] Jedes Monomer kann separat ein Molekül Thioredoxin binden.
-

Die Struktur von E coli ThxR-Dimer band Thioredoxin
-

Die Struktur von E coli ThxR mit markierten FAD- und NADPH-Prothesengruppen
Säugetier[edit]
Die TrxR-Struktur von Säugetieren ähnelt der E coli. Es enthält eine FAD- und NADPH-Bindungsdomäne sowie eine Schnittstelle zwischen zwei Monomeruntereinheiten. In ThxR von Säugetieren gibt es eine Insertion in die FAD-Bindungsdomäne zwischen zwei Alpha-Helices, die ein kleines Paar von Beta-Strängen bildet.[12] Das aktive Disulfid im Enzym befindet sich auf einer dieser Helices und somit befindet sich die aktive Disulfidbindung in der FAD-Domäne und nicht in der NADPH-Domäne wie in E coli und andere Prokaryoten.[12]
-

Struktur menschlicher ThxR FAD- und NADPH-Prothesengruppen
Mechanismus[edit]
E coli[edit]
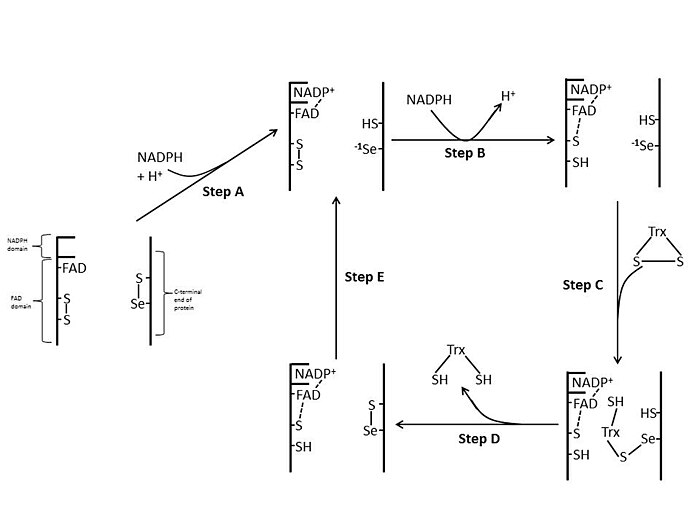
Im E coli Durch die räumliche Ausrichtung der FAD- und NADPH-Domänen sind die redoxaktiven Ringe von FAD und NADPH nicht nahe beieinander.[1] Wenn die FAD-Domäne von E coli wird um 66 Grad gedreht, wobei die NADPH-Domäne fixiert bleibt. Die beiden prothetischen Gruppen bewegen sich in engen Kontakt, so dass Elektronen von NADPH zu FAD und dann zur Disulfidbindung des aktiven Zentrums gelangen können.[1][15] Die konservierten Reste des aktiven Zentrums in E. coli sind -Cys-Ala-Thr-Cys-.[1]
Säugetier[edit]
Säugetier-TrxRs weisen eine viel höhere Sequenzhomologie mit Glutathionreduktase auf als E coli.[1] Die Cys-Reste des aktiven Zentrums in der FAD-Domäne und der gebundenen NADPH-Domäne befinden sich in unmittelbarer Nähe, wodurch die Notwendigkeit einer 66-Grad-Rotation für den Elektronentransfer in beseitigt wird E coli. Ein zusätzliches Merkmal des Säugetiermechanismus ist das Vorhandensein eines Selenocysteinrests am C-terminalen Ende des Proteins, das für die katalytische Aktivität erforderlich ist. Die konservierten Reste im aktiven Zentrum von Säugetieren sind -Cys-Val-Asn-Val-Gly-Cys-.[1]
Klinische Bedeutung[edit]
Krebsbehandlung[edit]
Da die Aktivität dieses Enzyms für das Zellwachstum und das Überleben wesentlich ist, ist es ein gutes Ziel für die Antitumor-Therapie. Darüber hinaus ist das Enzym bei verschiedenen Krebsarten, einschließlich des malignen Mesothelioms, hochreguliert.[16][17] Beispielsweise ist Motexafin Gadolinium (MGd) ein neues Chemotherapeutikum, das selektiv auf Tumorzellen abzielt und über die Hemmung von Thioredoxinreduktase und Ribonukleotidreduktase zum Zelltod und zur Apoptose führt.
Kardiomyopathie[edit]
Dilatierte Kardiomyopathie (DCM) ist eine häufige Diagnose bei Herzinsuffizienz. Thioredoxinreduktasen sind essentielle Proteine zur Regulierung des zellulären Redoxgleichgewichts und zur Minderung des Schadens, der durch reaktive Sauerstoffspezies verursacht wird, die durch oxidative Phosphorylierung in den Mitochondrien erzeugt werden. Die Inaktivierung von mitochondrialem TrxR2 bei Mäusen führt zu einer Ausdünnung der ventrikulären Herzwände und zum Tod des Neugeborenen.[10] Darüber hinaus finden sich zwei Mutationen im TrxR2-Gen bei Patienten, bei denen DCM diagnostiziert wurde, und nicht in einer Kontrollpopulation. Es wird angenommen, dass die pathologische Auswirkung dieser Mutationen eine beeinträchtigte Fähigkeit zur Kontrolle oxidativer Schäden in Herzmuskelzellen ist.[18]
Antibiotikum[edit]
Kürzlich wurden Untersuchungen durchgeführt, um zu zeigen, dass Thioredoxinreduktase mit niedrigem Molekulargewicht ein Ziel für neuartige Antibiotika (wie Auranofin oder Ebselen) sein könnte.[19]) Dies gilt insbesondere für Mycobacterium Haemophilumund könnte für antibiotikaresistente Bakterien verwendet werden.[20]
Verweise[edit]
- ^ ein b c d e f G Mustacich D, Powis G (Februar 2000). “Thioredoxinreduktase”. The Biochemical Journal. 346 Pt 1 (1): 1–8. doi:10.1042 / 0264-6021: 3460001. PMC 1220815. PMID 10657232.
- ^ Jordan A., Aslund F., Pontis E., Reichard P., Holmgren A. (Juli 1997). “Charakterisierung von Escherichia coli NrdH. Ein Glutaredoxin-ähnliches Protein mit einem Thioredoxin-ähnlichen Aktivitätsprofil”. Das Journal of Biological Chemistry. 272 (29): 18044–50. doi:10.1074 / jbc.272.29.18044. PMID 9218434.
- ^ Phulera S, Mande SC (Juni 2013). “Die Kristallstruktur von Mycobacterium tuberculosis NrdH bei 0,87 Å legt einen möglichen Aktivitätsmodus nahe”. Biochemie. 52 (23): 4056–65. doi:10.1021 / bi400191z. PMID 23675692.
- ^ Phulera S., Akif M., Sardesai AA, Mande SC (01.01.2014). “Redoxproteine von Mycobacterium tuberculosis”. Zeitschrift des Indian Institute of Science. 94 (1): 127–138. ISSN 0970-4140.
- ^ ein b c d Hirt RP, Müller S., Embley TM, Coombs GH (Juli 2002). “Die Vielfalt und Entwicklung der Thioredoxinreduktase: neue Perspektiven”. Trends in der Parasitologie. 18 (7): 302–8. doi:10.1016 / S1471-4922 (02) 02293-6. PMID 12379950.
- ^ ein b Holmgren A, Lu J (Mai 2010). “Thioredoxin und Thioredoxinreduktase: aktuelle Forschung unter besonderer Berücksichtigung menschlicher Krankheiten”. Biochemische und biophysikalische Forschungskommunikation. 396 (1): 120–4. doi:10.1016 / j.bbrc.2010.03.083. PMID 20494123.
- ^ Meyer Y., Buchanan BB, Vignols F., Reichheld JP (2009). “Thioredoxine und Glutaredoxine: verbindende Elemente in der Redoxbiologie”. Jahresrückblick auf die Genetik. 43: 335–67. doi:10.1146 / annurev-genet-102108-134201. PMID 19691428.
- ^ Lillig CH, Holmgren A (Januar 2007). “Thioredoxin und verwandte Moleküle – von der Biologie zu Gesundheit und Krankheit”. Antioxidantien & Redox-Signalisierung. 9 (1): 25–47. doi:10.1089 / ars.2007.9.25. PMID 17115886.
- ^ Arscott LD, Gromer S., Schirmer RH, Becker K., Williams CH (April 1997). “Der Mechanismus der Thioredoxinreduktase aus menschlicher Plazenta ähnelt den Mechanismen der Lipoamiddehydrogenase und der Glutathionreduktase und unterscheidet sich vom Mechanismus der Thioredoxinreduktase aus Escherichia coli.”. Verfahren der National Academy of Sciences der Vereinigten Staaten von Amerika. 94 (8): 3621–6. doi:10.1073 / pnas.94.8.3621. PMC 20490. PMID 9108027.
- ^ ein b Conrad M., Jakupoglu C., Moreno SG, Lippl S., Banjac A., Schneider M., Beck H., Hatzopoulos AK, Just U., Sinowatz F., Schmahl W., Chien KR, Wurst W., Bornkamm GW, Brielmeier M. (November 2004). “Wesentliche Rolle für die mitochondriale Thioredoxinreduktase bei Hämatopoese, Herzentwicklung und Herzfunktion”. Molekular- und Zellbiologie. 24 (21): 9414–23. doi:10.1128 / MCB.24.21.9414-9423.2004. PMC 522221. PMID 15485910.
- ^ ein b c Williams CH (Oktober 1995). “Mechanismus und Struktur der Thioredoxinreduktase aus Escherichia coli”. FASEB Journal. 9 (13): 1267–76. doi:10.1096 / fasebj.9.13.7557016. hdl:2027.42 / 154540. PMID 7557016. S2CID 26055087.
- ^ ein b Sandalova T., Zhong L., Lindqvist Y., Holmgren A., Schneider G. (August 2001). “Dreidimensionale Struktur einer Thioredoxinreduktase bei Säugetieren: Auswirkungen auf den Mechanismus und die Entwicklung eines Selenocystein-abhängigen Enzyms”. Verfahren der National Academy of Sciences der Vereinigten Staaten von Amerika. 98 (17): 9533–8. doi:10.1073 / pnas.171178698. PMC 55487. PMID 11481439.
- ^ Zhong L., Arnér ES, Holmgren A. (Mai 2000). Struktur und Mechanismus der Thioredoxinreduktase von Säugetieren: Das aktive Zentrum ist ein redoxaktives Selenolthiol / Selenenylsulfid, das aus der konservierten Cystein-Selenocystein-Sequenz gebildet wird.. Verfahren der National Academy of Sciences der Vereinigten Staaten von Amerika. 97 (11): 5854–9. doi:10.1073 / pnas.100114897. PMC 18523. PMID 10801974.
- ^ Becker K., Herold-Mende C., Park J. J., Lowe G., Schirmer RH (August 2001). “Humane Thioredoxinreduktase wird durch (2,2 ‘: 6’, 2 ” – Terpyridin) platin (II) -Komplexe wirksam gehemmt. Mögliche Implikationen für eine neuartige Antitumorstrategie”. Journal of Medicinal Chemistry. 44 (17): 2784–92. doi:10.1021 / jm001014i. PMID 11495589.
- ^ Lennon BW, Williams CH (August 1997). “Reduktive Halbreaktion von Thioredoxinreduktase aus Escherichia coli”. Biochemie. 36 (31): 9464–77. doi:10.1021 / bi970307j. PMID 9235991.
- ^ Nilsonne G., Sun X, Nyström C., Rundlöf AK, Potamitou Fernandes A., Björnstedt M., Dobra K. (September 2006). “Selenit induziert Apoptose in sarkomatoiden malignen Mesotheliomzellen durch oxidativen Stress”. Freie Radikale Biologie & Medizin. 41 (6): 874–85. doi:10.1016 / j.freeradbiomed.2006.04.031. PMID 16934670.
- ^ Kahlos K., Soini Y., Säily M., Koistinen P., Kakko S., Pääkkö P., Holmgren A., Kinnula VL (Mai 2001). “Hochregulierung von Thioredoxin und Thioredoxinreduktase beim malignen Pleuramesotheliom des Menschen”. Internationale Zeitschrift für Krebs. 95 (3): 198–204. doi:10.1002 / 1097-0215 (20010520) 95: 3<198::AID-IJC1034>3.0.CO; 2-F. PMID 11307155.
- ^ Geschwister D, Pfeufer A, Perisic T, Mannes AM, Fritz-Wolf K, Unwin S, Sünder MF, Gieger C, Glöckner CJ, Wichmann HE, Kremmer E, Schäfer Z, Walch A, Hinterseer M, Näbauer M, Kääb S, Kastrati A., Schömig A., Meitinger T., Bornkamm GW, Conrad M., von Beckerath N. (Mai 2011). “Mutationen im mitochondrialen Thioredoxinreduktase-Gen TXNRD2 verursachen eine erweiterte Kardiomyopathie.”. Europäisches Herzjournal. 32 (9): 1121–33. doi:10.1093 / eurheartj / ehq507. PMID 21247928.
- ^ Marshall AC, Kidd SE, Lamont-Friedrich SJ, Arentz G., Hoffmann P., Coad BR, Bruning JB (März 2019). “Aspergillus fumigatus Thioredoxin Reductase”. Antimikrobielle Mittel und Chemotherapie. 63 (3). doi:10.1128 / AAC.02281-18. PMC 6395915. PMID 30642940.
- ^ Harbut MB, Vilchèze C., Luo X, Hensler ME, Guo H., Yang B. et al. (April 2015). “Auranofin übt bakterizide Breitbandaktivitäten aus, indem es auf die Thiol-Redox-Homöostase abzielt.”. Verfahren der National Academy of Sciences der Vereinigten Staaten von Amerika. 112 (14): 4453–8. doi:10.1073 / pnas.1504022112. PMC 4394260. PMID 25831516.
Externe Links[edit]
Recent Comments