Flugsaurier – Wikipedia
Fliegende Reptilien der ausgestorbenen Gruppe oder Ordnung Pterosauria
Flugsaurier (;[4][5] aus dem Griechischen Pteron und Sauros, was “Flügeleidechse” bedeutet) waren fliegende Reptilien der ausgestorbenen Gruppe oder Ordnung Flugsaurier. Sie existierten während des größten Teils des Mesozoikums: von der späten Trias bis zum Ende der Kreidezeit (vor 228 bis 66 Millionen Jahren)[6]). Flugsaurier sind die frühesten Wirbeltiere, von denen bekannt ist, dass sie einen Motorflug entwickelt haben. Ihre Flügel wurden von einer Membran aus Haut, Muskeln und anderen Geweben gebildet, die sich von den Knöcheln bis zu einem dramatisch verlängerten vierten Finger erstreckten.[7]
Es gab zwei Haupttypen von Flugsauriern. Basale Flugsaurier (auch als “nicht-pterodaktyloide Flugsaurier” oder “Rhamphorhynchoide” bezeichnet) waren kleinere Tiere mit voll gezahnten Kiefern und typischerweise langen Schwänzen. Ihre breiten Flügelmembranen umfassten wahrscheinlich die Hinterbeine und verbanden sie. Am Boden hätten sie eine unangenehme Haltung gehabt, aber ihre Gelenkanatomie und ihre starken Krallen hätten sie zu effektiven Kletterern gemacht, und sie hätten möglicherweise in Bäumen gelebt. Basale Flugsaurier waren Insektenfresser oder Raubtiere kleiner Wirbeltiere. Später entwickelten Pterosaurier (Pterodactyloide) viele Größen, Formen und Lebensstile. Pterodactyloide hatten schmalere Flügel mit freien Hinterbeinen, stark reduzierten Schwänzen und langen Hälsen mit großen Köpfen. Auf dem Boden liefen Pterodaktyloide auf allen vier Gliedmaßen mit aufrechter Haltung gut, standen Plantigrade auf den Hinterfüßen und falteten den Flügelfinger nach oben, um auf der dreifingrigen “Hand” zu gehen. Sie konnten vom Boden abheben, und fossile Wege zeigen, dass zumindest einige Arten rennen und waten oder schwimmen konnten.[8] Ihre Kiefer hatten geile Schnäbel und einigen Gruppen fehlten Zähne. Einige Gruppen entwickelten ausgefeilte Kopfkämme mit sexuellem Dimorphismus.
Flugsaurier trugen Mäntel aus haarartigen Filamenten, die als Pyknofasern bekannt sind und ihre Körper und Teile ihrer Flügel bedeckten. Pyknofasern wuchsen in verschiedenen Formen, von einfachen Filamenten bis hin zu verzweigten Federn. Diese sind möglicherweise homolog zu den Daunenfedern, die sowohl bei Vogel- als auch bei einigen Nicht-Vogel-Dinosauriern gefunden wurden, was darauf hindeutet, dass sich frühe Federn bei dem gemeinsamen Vorfahren von Flugsauriern und Dinosauriern entwickelt haben, möglicherweise als Isolierung.[9] Im Leben hätten Flugsaurier glatte oder flauschige Mäntel gehabt, die nicht wie Vogelfedern aussahen. Sie waren warmblütige (endotherme) aktive Tiere. Das Atmungssystem hatte eine effiziente unidirektionale “Durchfluss” -Atmung unter Verwendung von Luftsäcken, die ihre Knochen in extremem Maße aushöhlen. Flugsaurier umfassten eine breite Palette von Erwachsenengrößen, von den sehr kleinen Anurognathiden bis zu den größten bekannten fliegenden Kreaturen aller Zeiten, einschließlich Quetzalcoatlus und Hatzegopteryx,[10][11][12] die Flügelspannweiten von mindestens neun Metern erreichte. Die Kombination aus Endothermie, guter Sauerstoffversorgung und starken Muskeln machte Flugsaurier zu kraftvollen und fähigen Fliegern.
Flugsaurier werden von den populären Medien oder der Öffentlichkeit oft als “fliegende Dinosaurier” bezeichnet, aber Dinosaurier werden als Nachkommen des letzten gemeinsamen Vorfahren der Saurischia und Ornithischia definiert, was die Pterosaurier ausschließt.[13] Flugsaurier sind jedoch enger mit Vögeln und anderen Dinosauriern verwandt als mit Krokodilen oder anderen lebenden Reptilien, obwohl sie keine Vorfahren der Vögel sind. Flugsaurier werden auch umgangssprachlich als bezeichnet Pterodaktylenvor allem in der Fiktion und von Journalisten.[14] Technisch gesehen jedoch Pterodaktylus bezieht sich nur auf Mitglieder der Gattung Pterodactylusund allgemeiner an Mitglieder der Unterordnung Pterodactyloidea der Pterosaurier.[15]
Flugsaurier hatten eine Vielzahl von Lebensstilen. Traditionell als Fischfresser angesehen, umfasst die Gruppe heute Jäger von Landtieren, Insektenfressern, Obstfressern und sogar Raubtieren anderer Flugsaurier. Sie vermehrten sich durch Eier, von denen einige Fossilien entdeckt wurden.
Beschreibung[edit]
Die Anatomie der Flugsaurier wurde durch die Anpassung an die Flucht gegenüber ihren Reptilienvorfahren stark verändert. Flugsaurierknochen waren hohl und luftgefüllt wie die von Vögeln. Dies lieferte eine höhere Muskelbefestigungsfläche für ein gegebenes Skelettgewicht. Die Knochenwände waren oft hauchdünn. Sie hatten ein großes Brustbein mit Kiel für die Flugmuskulatur und ein vergrößertes Gehirn, das das komplexe Flugverhalten koordinieren konnte.[16] Flugsaurierskelette zeigen oft eine beträchtliche Verschmelzung. Im Schädel verschwanden die Nähte zwischen den Elementen. Bei einigen späteren Flugsauriern verschmolz das Rückgrat über den Schultern zu einer als Notarium bekannten Struktur, die dazu diente, den Oberkörper während des Fluges zu versteifen und das Schulterblatt stabil zu stützen. Ebenso könnten die Sakralwirbel ein einziges Synsacrum bilden, während auch die Beckenknochen verschmolzen.
Basale Flugsaurier umfassen die Kladen Dimorphodontidae (Dimorphodon), Campylognathididae (Eudimorphodon, Campyognathoides) und Rhamphorhynchidae (Rhamphorhynchus, Scaphognathus).
Pterodactyloide umfassen die Kladen Ornithocheiroidea (Istiodactylus, Ornithocheirus, Pteranodon), Ctenochasmatoidea (Ctenochasma, Pterodactylus), Dsungaripteroidea (Germanodactylus, Dsungaripterus) und Azhdarchoidea (Tapejara, Tupuxuara, Quetzalcoatlus).
Die beiden Gruppen überlappten sich zeitlich, aber die frühesten Flugsaurier im Fossilienbestand sind basale Flugsaurier, und die neuesten Flugsaurier sind Pterodaktyloide.[17]
Die Position der Klade Anurognathidae (Anurognathus, Jeholopterus, Vesperopterylus) wird diskutiert.[18] Anurognathiden (Pterosaurier mit Froschkopf) waren hochspezialisiert. Kleine Flyer mit verkürzten Kiefern und großem Spalt, einige hatten große Augen, die auf nächtliche oder kreppartige Gewohnheiten hindeuteten, Mundborsten und Füße, die zum Festhalten geeignet waren. Parallele Anpassungen sind bei Vögeln und Fledermäusen zu beobachten, die im Flug Insekten jagen.
Größe[edit]
Flugsaurier hatten eine große Auswahl an Größen. Im Allgemeinen waren sie ziemlich groß. Selbst die kleinste Art hatte eine Flügelspannweite von nicht weniger als 25 Zentimetern.[10] Die größten Formen stellen die größten bekannten Tiere dar, die jemals geflogen sind, mit Flügelspannweiten von bis zu 10 bis 11 Metern.[19]
Im Stehen könnten solche Riesen die Höhe einer modernen Giraffe erreichen. Traditionell wurde angenommen, dass Flugsaurier im Verhältnis zu ihrer Größe extrem leicht waren. Später wurde verstanden, dass dies unrealistisch niedrige Dichten ihrer Weichteile implizieren würde. Einige moderne Schätzungen extrapolieren daher für die größten Arten ein Gewicht von bis zu 250 Kilogramm.
Schädel, Zähne und Kämme[edit]

Im Vergleich zu den anderen Wirbeltierfluggruppen, den Vögeln und Fledermäusen, waren die Schädel der Flugsaurier typischerweise ziemlich groß. Die meisten Flugsaurierschädel hatten verlängerte Kiefer. Ihre Schädelknochen neigen dazu, bei erwachsenen Personen verwachsen zu sein. Frühe Flugsaurier hatten oft heterodontische Zähne mit unterschiedlichem Körperbau, und einige hatten noch Zähne am Gaumen. In späteren Gruppen wurden die Zähne meist konisch.[22] Die Vorderzähne waren oft länger und bildeten einen “Beutegreifer” in quer ausgedehnten Kieferspitzen, aber Größe und Position waren bei den Arten sehr unterschiedlich. Mit der abgeleiteten Pterodactyloidea wurden die Schädel noch länglicher und übertrafen manchmal den kombinierten Hals und Rumpf in der Länge. Dies wurde durch eine Dehnung und Verschmelzung des vorderen Schnauzenknochens, der Prämaxilla, mit dem Oberkieferknochen, dem Oberkiefer, verursacht. Im Gegensatz zu den meisten Archosauriern verschmolzen die nasalen und antorbitalen Öffnungen von Pterodaktyloid-Pterosauriern zu einer einzigen großen Öffnung, der so genannten nasoantorbitales fenestra. Dieses Merkmal hat sich wahrscheinlich entwickelt, um den Schädel für den Flug aufzuhellen.[22] Im Gegensatz dazu zogen sich die Knochen hinter der Augenhöhle zusammen und drehten sich, wobei der hintere Schädel stark geneigt und das Kiefergelenk nach vorne gebracht wurde. Die Gehirnschale war für Reptilien relativ groß.
In einigen Fällen ist versteinertes keratinhaltiges Schnabelgewebe erhalten geblieben, obwohl der Schnabel in Zahnform klein und auf die Kieferspitzen beschränkt ist und die Zähne nicht betrifft.[27] Einige fortgeschrittene Schnabelformen waren zahnlos, wie die Pteranodontidae und Azhdarchidae, und hatten größere, umfangreichere und vogelähnlichere Schnäbel.[22] Einige Gruppen hatten spezielle Zahnformen. Die Istiodactylidae hatten zurückgebogene Zähne, um Fleisch zu essen. Ctenochasmatidae verwendeten Kämme mit zahlreichen nadelartigen Zähnen zur Filterfütterung; Pterodaustro könnte über tausend borstenartige Zähne haben. Dsungaripteridae bedeckten ihre Zähne mit Kieferknochengewebe für eine Quetschfunktion. Wenn Zähne vorhanden waren, wurden sie in separate Zahnpfannen gelegt. Ersatzzähne wurden hinter und nicht unter den älteren Zähnen erzeugt.

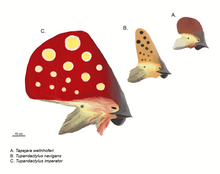
Das öffentliche Bild von Flugsauriern wird durch ihre kunstvollen Kopfkämme definiert. Dies wurde durch das markante nach hinten weisende Wappen des Bekannten beeinflusst Pteranodon. Die Hauptpositionen solcher Kämme sind die Vorderseite der Schnauze als Auswuchs der Prämaxillen oder die Rückseite des Schädels als Verlängerung der Scheitelknochen. In diesem Fall wird sie als “supraoccipitaler Kamm” bezeichnet. Vordere und hintere Kämme können gleichzeitig vorhanden sein und können zu einer einzigen größeren Struktur verschmolzen sein, von der die Tapejaridae die expansivste zeigen. Nyctosaurus trug ein bizarres geweihartiges Wappen. Die Kämme waren quer nur wenige Millimeter dünn. Die knöcherne Kammbasis würde typischerweise durch keratinöses oder anderes Weichgewebe verlängert.
Seit den 1990er Jahren haben neue Entdeckungen und eine gründlichere Untersuchung alter Exemplare gezeigt, dass Kämme unter Flugsauriern weitaus weiter verbreitet sind als bisher angenommen. Dass sie durch Keratin erweitert wurden oder vollständig aus Keratin bestehen, das nicht leicht versteinert, hatte frühere Forschungen in die Irre geführt.[29] Zum Pterorhynchus und PterodactylusDas wahre Ausmaß dieser Kämme wurde nur mithilfe der Ultraviolettfotografie entdeckt.[27][30] Während fossile Kämme früher auf die fortgeschrittenere Pterodactyloidea beschränkt waren, Pterorhynchus und Austriadactylus zeigen, dass sogar einige frühe Flugsaurier sie besaßen.[29]
Wie die Oberkiefer waren auch die gepaarten Unterkiefer der Flugsaurier sehr länglich. In fortgeschrittenen Formen waren sie tendenziell kürzer als der obere Schädel, da sich das Kiefergelenk in einer weiter vorne liegenden Position befand. Die vorderen Unterkieferknochen, die Zähne oder Ossa Dentaliawurden an der Spitze fest zu einer zentralen Symphyse verschmolzen. Dadurch funktionierten die Unterkiefer als ein einziges verbundenes Ganzes, der Unterkiefer. Die Symphyse war oft quer und lang sehr dünn und machte einen beträchtlichen Teil der Kieferlänge aus, bis zu 60%. Wenn ein Kamm auf der Schnauze vorhanden war, könnte die Symphyse einen passenden Unterkieferkamm aufweisen, der nach unten ragt. Zahnarten trugen auch Zähne in ihren Zähnen. Der Unterkiefer öffnete und schloss sich in einer einfachen vertikalen oder “orthalen” Auf- und Abbewegung.
Wirbelsäule[edit]

Die Wirbelsäule der Flugsaurier war zwischen vierunddreißig und siebzig Wirbeln nummeriert. Die Wirbel vor dem Schwanz waren “prokoel”: Der Cotylus (Vorderseite des Wirbelkörpers) war konkav und passte eine konvexe Verlängerung an der Rückseite des vorhergehenden Wirbels, des Kondylus, hinein. Fortgeschrittene Flugsaurier besitzen einzigartige spezielle Prozesse, die neben ihrem Kondylus und Cotylus, den Exapophysen, hervorstehen.[32] und der Cotyle kann auch einen kleinen Stift auf seiner Mittellinie besitzen, der als Hypapophyse bezeichnet wird.

Der Hals der Flugsaurier war relativ lang und gerade. Bei Pterodaktyloiden ist der Hals typischerweise länger als der Oberkörper. Diese Länge wird nicht durch eine Zunahme der Anzahl der Wirbel verursacht, die ausnahmslos sieben beträgt. Einige Forscher schließen zwei vorübergehende “Cervicodorsale” ein, die die Zahl auf neun bringen. Stattdessen wurden die Wirbel selbst länglicher, bis zu achtmal länger als breit. Trotzdem war der Gebärmutterhals breiter als hoch, was eine bessere vertikale als horizontale Halsbeweglichkeit impliziert. Pterodactyloide haben alle Halsrippen verloren. Pterosaurierhälse waren wahrscheinlich ziemlich dick und muskulös, besonders vertikal.
Der Oberkörper war relativ kurz und eiförmig. Die Wirbel im Rücken der Flugsaurier könnten ursprünglich achtzehn gewesen sein. Bei fortgeschrittenen Arten neigte eine wachsende Anzahl davon dazu, in das Kreuzbein eingebaut zu werden. Solche Arten zeigen auch häufig eine Verschmelzung der vorderen Rückenwirbel zu einem starren Ganzen, das nach einer vergleichbaren Struktur bei Vögeln als Notarium bezeichnet wird. Dies war eine Anpassung, um den Kräften standzuhalten, die durch das Schlagen der Flügel verursacht wurden. Das Notarium umfasste je nach Art drei bis sieben Wirbel, aber auch das individuelle Alter. Diese Wirbel könnten durch Sehnen oder eine Verschmelzung ihrer Nervenstacheln zu einer “supraneuralen Platte” verbunden sein. Ihre Rippen würden auch fest mit dem Notarium verschmolzen sein. Im Allgemeinen sind die Rippen doppelköpfig. Das Kreuzbein bestand aus drei bis zehn Sakralwirbeln. Auch sie könnten über eine supraneurale Platte verbunden werden, die jedoch das Notarium nicht berührt.

Die Schwänze der Flugsaurier waren immer ziemlich schlank. Dies bedeutet, dass der Caudofemoralis-Retraktor-Muskel, der in den meisten basalen Archosaurien die Hauptantriebskraft für das Hinterbein darstellt, relativ unwichtig war. Die Schwanzwirbel waren amphikoelös, die Wirbelkörper an beiden Enden waren konkav. Frühe Arten hatten lange Schwänze, die bis zu fünfzig Schwanzwirbel enthielten, wobei die mittleren durch langgestreckte Artikulationsprozesse, Zygapophysen und Chevrons versteift waren. Solche Schwänze fungierten als Ruder und endeten manchmal hinten in einer vertikalen rautenförmigen oder ovalen Schaufel. Bei Pterodaktyloiden waren die Schwänze stark reduziert und nie versteift, wobei einige Arten nur zehn Wirbel zählten.
Schultergürtel[edit]
Der Schultergürtel war eine starke Struktur, die die Kräfte des flatternden Fluges auf den Thorax übertrug. Es war wahrscheinlich von dicken Muskelschichten bedeckt. Der obere Knochen, das Schulterblatt, war eine gerade Stange. Es war mit einem unteren Knochen verbunden, dem Coracoid, das bei Flugsauriern relativ lang ist. Bei fortgeschrittenen Arten war ihr kombiniertes Ganzes, das Scapulocoracoid, fast vertikal ausgerichtet. Das Schulterblatt passte in diesem Fall in eine Aussparung an der Seite des Notariums, während das Coracoid ebenfalls mit dem Brustbein verbunden war. Auf diese Weise bildeten beide Seiten zusammen eine starre geschlossene Schleife, die erheblichen Kräften standhalten konnte. Eine Besonderheit war, dass die Brustbeinverbindungen der Coracoide oft asymmetrisch waren, wobei ein Coracoid vor dem anderen angebracht war. Bei fortgeschrittenen Arten hatte sich das Schultergelenk vom Schulterblatt zum Coracoid bewegt. Das Gelenk war sattelförmig und ermöglichte eine beträchtliche Bewegung des Flügels. Es war seitwärts und etwas nach oben gerichtet.
Das Brustbein, gebildet durch verschmolzenes Paar Sternawar breit. Es hatte nur einen flachen Kiel. Über sternale Rippen war es an seinen Seiten an den Rückenrippen befestigt. Auf seiner Rückseite befand sich eine Reihe von Bauchrippen oder Gastralien, die den gesamten Bauch bedeckten. Nach vorne ein langer Punkt, der Cristospinaschräg nach oben ragen. Die Hinterkante des Brustbeins war der tiefste Punkt des Thorax. Schlüsselbeine oder Interklavikel fehlten vollständig.
Flügel[edit]
Flugsaurierflügel wurden durch Knochen und Membranen der Haut und anderer Gewebe gebildet. Die Primärmembranen sind am extrem langen vierten Finger jedes Arms befestigt und erstrecken sich entlang der Körperseiten. Wo sie endeten, war sehr umstritten, aber seit den 1990er Jahren wurden ein Dutzend Exemplare mit konserviertem Weichgewebe gefunden, die zu zeigen scheinen, dass sie an den Knöcheln haften. Die genaue Krümmung der Hinterkante ist jedoch immer noch nicht eindeutig.

Während sie historisch als einfache ledrige Strukturen aus Haut angesehen wurden, hat die Forschung seitdem gezeigt, dass die Flügelmembranen von Flugsauriern hochkomplexe dynamische Strukturen waren, die für einen aktiven Flugstil geeignet waren. Die äußeren Flügel (von der Spitze bis zum Ellbogen) wurden durch eng beieinander liegende Fasern verstärkt Actinofibrillen.[45] Die Actinofibrillen selbst bestanden aus drei verschiedenen Schichten im Flügel, die ein Kreuzmuster bildeten, wenn sie einander überlagert wurden. Die Funktion der Actinofibrillen ist unbekannt, ebenso wie das genaue Material, aus dem sie hergestellt wurden. Abhängig von ihrer genauen Zusammensetzung (Keratin, Muskel, elastische Strukturen usw.) können sie im äußeren Teil des Flügels Versteifungs- oder Verstärkungsmittel gewesen sein.[46] Die Flügelmembranen enthielten auch eine dünne Muskelschicht, faseriges Gewebe und ein einzigartiges, komplexes Kreislaufsystem, das Blutgefäße schleift.[29] Die Kombination von Aktinofibrillen und Muskelschichten kann es dem Tier ermöglicht haben, die Flügelschwäche und den Sturz einzustellen.
Wie Hohlräume in den Flügelknochen größerer Arten und in mindestens einer Probe konserviertes Weichgewebe zeigen, erweiterten einige Flugsaurier ihr System von Atemluftsäcken in die Flügelmembran.[47]
Teile des Flügels[edit]

Die Pterosaurier-Flügelmembran ist in drei Grundeinheiten unterteilt. Der erste, genannt der Propatagium (“Vordermembran”) war der vorderste Teil des Flügels und wurde zwischen Handgelenk und Schulter befestigt, wodurch die “Vorderkante” während des Fluges entstand. Das Brachiopatagium (“Armmembran”) war der Hauptbestandteil des Flügels und erstreckte sich vom stark verlängerten vierten Finger der Hand bis zu den Hinterbeinen. Schließlich hatten zumindest einige Flugsauriergruppen eine Membran, die sich zwischen den Beinen erstreckte und möglicherweise mit dem Schwanz verbunden war oder diesen einbaute Uropatagium;; Das Ausmaß dieser Membran ist nicht sicher, wie Studien an Sordes scheinen darauf hinzudeuten, dass es einfach die Beine verband, aber nicht den Schwanz einbezog (was es a Cruropatagium). Eine verbreitete Interpretation ist, dass nicht-pterodaktyloide Pterosaurier ein breiteres Uro / Cruropatagium hatten, das zwischen ihren langen fünften Zehen gespannt war, wobei Pterodaktyloide, denen solche Zehen fehlten, nur Membranen hatten, die entlang der Beine liefen.
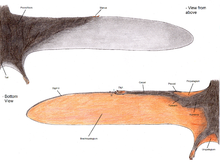
Paläontologen haben erhebliche Auseinandersetzungen darüber geführt, ob und wo die Hauptflügelmembranen (Brachiopatagie) an den Hinterbeinen haften. Fossilien des Rhamphorhynchoid Sordes,[50] der Anurognathid Jeholopterus,[51] und ein Pterodactyloid aus der Santana-Formation scheint zu zeigen, dass sich die Flügelmembran zumindest bei einigen Arten an den Hinterbeinen festsetzte.[52] Moderne Fledermäuse und fliegende Eichhörnchen weisen jedoch erhebliche Unterschiede im Ausmaß ihrer Flügelmembranen auf, und es ist möglich, dass wie diese Gruppen verschiedene Arten von Flugsauriern unterschiedliche Flügeldesigns hatten. Tatsächlich zeigt die Analyse der Proportionen der Pterosaurierglieder, dass es erhebliche Unterschiede gab, die möglicherweise eine Vielzahl von Flügelplänen widerspiegeln.[53]
Die knöchernen Elemente des Arms bildeten einen Mechanismus zum Stützen und Ausfahren des Flügels. In der Nähe des Körpers ist der Humerus oder Oberarmknochen kurz, aber kräftig gebaut. Es hat einen großen Deltopektoralkamm, an dem die Hauptflugmuskeln befestigt sind. Trotz der erheblichen Kräfte, die auf ihn ausgeübt werden, ist der Humerus innen hohl oder pneumatisiert, verstärkt durch Knochenstreben. Die langen Knochen des Unterarms, die Ulna und der Radius sind viel länger als der Humerus. Sie waren wahrscheinlich nicht zur Pronation fähig.
Ein für Pterosaurier einzigartiger Knochen, bekannt als Pteroid, der mit dem Handgelenk verbunden ist und dabei hilft, die vordere Membran (das Propatagium) zwischen Handgelenk und Schulter zu stützen. Hinweise auf ein Gurtband zwischen den drei freien Fingern des Pterosaurier-Vorderbeins deuten darauf hin, dass diese vordere Membran möglicherweise umfangreicher war als die einfache Verbindung zwischen Pteroid und Schulter, die traditionell bei Lebensrestaurationen dargestellt wird.[29] Die Position des Pteroidknochens selbst war umstritten. Einige Wissenschaftler, insbesondere Matthew Wilkinson, haben argumentiert, dass der Pteroid nach vorne zeigte, die vordere Membran verlängerte und sie als einstellbare Klappe fungieren ließ.[57] Diese Ansicht wurde in einem Artikel von Chris Bennett aus dem Jahr 2007 widerlegt, der zeigte, dass der Pteroid nicht wie zuvor angenommen artikuliert war und nicht nach vorne hätte zeigen können, sondern wie traditionell interpretiert nach innen auf den Körper gerichtet war.[58] David Peters schlug 2009 vor, dass das Pteroid, das mit dem “Sattel” der Radiale (proximaler Synkarpal) artikuliert ist, und sowohl das Pteroid als auch das präaxiale Karpal zentral zentriert sind.[59] Dies scheint durch Exemplare von bestätigt zu werden Changchengopterus pani und Darwinopterus linglongtaensis zeigt den Pteroid in Artikulation mit dem proximalen Synkarpal.[60][61]
Das Pterosaurier-Handgelenk besteht aus zwei inneren (proximal, an der Seite der langen Knochen des Arms) und vier äußeren (distal, an der Seite der Hand) Karpalen (Handgelenkknochen), mit Ausnahme des Pteroidknochens, der selbst a sein kann modifizierter distaler Karpal. Die proximalen Karpalen sind in reifen Exemplaren zu einem “Synkarpal” verschmolzen, während drei der distalen Karpalen zu einem distalen Synkarpal verschmelzen. Der verbleibende distale Karpal, hier als medialer Karpal bezeichnet, der aber auch als distaler lateraler oder präaxialer Karpal bezeichnet wird, artikuliert auf einer vertikal länglichen bikonvexen Facette an der Vorderfläche des distalen Synkarpals. Die mediale Handwurzel trägt eine tiefe konkave Fovea, die sich nach Wilkinson anterior, ventral und etwas medial öffnet, in der sich der Pteroid artikuliert.[62]
In abgeleiteten Pterodaktyloiden wie Pteranodonten und Azhdarchoiden sind die Metakarpalen I-III klein und verbinden sich nicht mit dem Karpus, sondern hängen in Kontakt mit dem vierten Metakarpal. Bei diesen abgeleiteten Arten ist der vierte Mittelhandknochen enorm verlängert, was typischerweise der Länge der langen Knochen des Unterarms entspricht oder diese übersteigt. Der fünfte Mittelhandknochen war verloren gegangen. Bei allen Arten sind der erste bis dritte Finger viel kleiner als der vierte, der “Flügelfinger”, und enthalten zwei, drei bzw. vier Phalangen. Die kleineren Finger sind gekratzt, wobei die unguale Größe zwischen den Arten variiert. Bei Nyctosauriden sind die Vorderbeinziffern neben dem Flügelfinger vollständig verloren gegangen. Der Flügelfinger macht etwa die Hälfte oder mehr der gesamten Flügellänge aus. Es besteht normalerweise aus vier Phalangen. Ihre relativen Längen variieren tendenziell zwischen den Arten, was häufig zur Unterscheidung verwandter Formen verwendet wurde. Die vierte Phalanx ist normalerweise die kürzeste. Es fehlt eine Klaue und wurde von Nyctosauriden vollständig verloren. Es ist nach hinten gebogen, was zu einer abgerundeten Flügelspitze führt, die den induzierten Widerstand verringert. Der Flügelfinger ist ebenfalls etwas nach unten gebogen.
Im Stehen ruhten Flugsaurier wahrscheinlich auf ihren Mittelhandknochen, wobei der äußere Flügel nach hinten gefaltet war. In dieser Position wurden die “vorderen” Seiten der Mittelhandknochen nach hinten gedreht. Dies würde die kleineren Finger schräg nach hinten zeigen. Laut Bennett würde dies bedeuten, dass der Flügelfinger, der den größten Bogen eines Flügelelements bis zu 175 ° beschreiben kann, nicht durch Beugung, sondern durch eine extreme Streckung gefaltet wurde. Der Flügel wurde automatisch gefaltet, wenn der Ellbogen gebeugt wurde.
Becken[edit]

Das Becken der Flugsaurier war im Vergleich zum gesamten Körper mäßig groß. Oft waren die drei Beckenknochen verwachsen. Das Ilium war lang und niedrig, und seine vorderen und hinteren Klingen ragten horizontal über die Ränder der unteren Beckenknochen hinaus. Trotz dieser Länge weist die stabförmige Form dieser Prozesse darauf hin, dass die an ihnen befestigten Hinterbeinmuskeln nur eine begrenzte Kraft hatten. Der in der Seitenansicht schmale Schambein verschmolz mit dem breiten Ischium zu einer ischiopubischen Klinge. Manchmal waren auch die Klingen beider Seiten verschmolzen, wodurch das Becken von unten geschlossen und der Beckenkanal gebildet wurde. Das Hüftgelenk war nicht perforiert und ermöglichte eine beträchtliche Beweglichkeit des Beins. Es war schräg nach oben gerichtet, wodurch eine perfekt vertikale Position des Beins verhindert wurde.
Die Vorderseite der Schambeinknochen ist mit einer einzigartigen Struktur gegliedert, den gepaarten präpubischen Knochen. Zusammen bildeten diese eine Spitze, die den hinteren Bauch zwischen dem Becken und den Bauchrippen bedeckte. Die vertikale Beweglichkeit dieses Elements deutet auf eine Atmungsfunktion hin, die die relative Steifheit der Brusthöhle kompensiert.
Hintere Gliedmaßen[edit]
Die Hinterbeine der Flugsaurier waren stark gebaut, aber im Verhältnis zu ihren Flügelspannweiten kleiner als die der Vögel. Sie waren lang im Vergleich zur Rumpflänge. Der Oberschenkelknochen war ziemlich gerade, wobei der Kopf nur einen kleinen Winkel zum Schaft bildete. Dies impliziert, dass die Beine nicht vertikal unter dem Körper gehalten wurden, sondern sich etwas ausbreiteten. Das Schienbein wurde oft mit den oberen Knöchelknochen zu einem Tibiotarsus verschmolzen, der länger als der Oberschenkelknochen war. Es könnte beim Gehen eine vertikale Position erreichen. Der Wadenknochen war tendenziell schlank, insbesondere an seinem unteren Ende, das in fortgeschrittenen Formen den Knöchel nicht erreichte, und reduzierte manchmal die Gesamtlänge auf ein Drittel. Typischerweise wurde es mit dem Schienbein verschmolzen. Der Knöchel war ein einfaches “mesotarsales” Scharnier. Der ziemlich lange und schlanke Mittelfuß war immer bis zu einem gewissen Grad gespreizt. Der Fuß war plantigrade, was bedeutet, dass während des Gehzyklus die Sohle des Mittelfußes auf den Boden gedrückt wurde.
In Bezug auf die Form der fünften Ziffer gab es einen deutlichen Unterschied zwischen frühen Flugsauriern und fortgeschrittenen Arten. Ursprünglich war der fünfte Mittelfußknochen robust und nicht sehr verkürzt. Es war in einer höheren Position als die anderen Mittelfußknochen mit dem Knöchel verbunden. Es trug einen langen und oft gebogenen, beweglichen klauenlosen fünften Zeh, der aus zwei Phalangen bestand. Die Funktion dieses Elements war rätselhaft. Früher dachte man, die Tiere schliefen kopfüber wie Fledermäuse, hingen an Ästen und benutzten die fünften Zehen als Haken. Eine andere Hypothese besagte, dass sie die Brachiopatagie dehnten, aber bei artikulierten Fossilien sind die fünften Ziffern immer zum Schwanz hin gebogen. Später wurde es populär anzunehmen, dass diese Zehen ein Uropatagium oder Cruropatagium zwischen ihnen verlängerten. Da sich die fünften Zehen außerhalb der Füße befanden, wäre eine solche Konfiguration nur möglich gewesen, wenn diese im Flug ihre Fronten nach außen gedreht hätten. Eine solche Rotation könnte durch eine Abduktion des Oberschenkelknochens verursacht werden, was bedeutet, dass die Beine gespreizt würden. Dies würde auch die Füße in eine vertikale Position bringen. Sie könnten dann als Ruder fungieren, um das Gieren zu kontrollieren. Einige Proben zeigen Membranen zwischen den Zehen,[69] Dadurch können sie als Flugsteuerflächen fungieren. Das (cr) Uroptagium würde die Tonhöhe steuern. Beim Gehen können sich die Zehen nach oben beugen, um die Membran vom Boden abzuheben. Bei Pterodactyloidea war der fünfte Mittelfuß stark reduziert und der fünfte Zeh, falls vorhanden, kaum mehr als ein Stummel. Dies deutet darauf hin, dass ihre Membranen gespalten waren, was die Flugmanövrierfähigkeit erhöhte.
Die ersten bis vierten Zehen waren lang. Sie hatten zwei, drei, vier und fünf Phalangen. Oft war der dritte Zeh am längsten; manchmal der vierte. Flache Gelenke weisen auf eine eingeschränkte Beweglichkeit hin. Diese Zehen waren gekratzt, aber die Krallen waren kleiner als die Handkrallen.
Weichteile[edit]
Die seltenen Bedingungen, die die Fossilisierung des Flugsauriers ermöglichten, bleiben bestehen, manchmal auch erhaltene Weichteile. Die moderne Synchrotron- oder Ultraviolettlichtfotografie hat viele Spuren gezeigt, die mit bloßem Auge nicht sichtbar sind. Diese werden oft ungenau als “Eindrücke” bezeichnet, bestehen jedoch meist aus Versteinerungen, natürlichen Abgüssen und Transformationen des Originalmaterials. Sie können Hornkämme, Schnäbel oder Klauenscheiden sowie die verschiedenen Flugmembranen umfassen. Ausnahmsweise blieben die Muskeln erhalten. Hautflecken zeigen kleine runde, nicht überlappende Schuppen an den Fußsohlen, den Knöcheln und den Enden der Mittelhandknochen. Sie bedeckten Polster, die den Aufprall des Gehens abfederten. Schuppen sind aus anderen Körperteilen unbekannt.
Pyknofasern[edit]

Die meisten oder alle Flugsaurier hatten haarartige Filamente, die als Pyknofasern bekannt sind, an Kopf und Rumpf. Der Begriff “Pyknofaser”, der “dichtes Filament” bedeutet, wurde 2009 vom Paläontologen Alexander Kellner und Kollegen geprägt.[46] Pyknofasern waren einzigartige Strukturen, die Säugetierhaaren ähnlich, aber nicht homolog waren (einen gemeinsamen Ursprung haben), ein Beispiel für konvergente Evolution.[50] Ein unscharfes Integument wurde zuerst von einer Probe von berichtet Scaphognathus crassirostris 1831 von Georg Augustus Goldfuss,[76] war aber weithin angezweifelt worden. Seit den 1990er Jahren haben Flugsaurierfunde sowie histologische und ultraviolette Untersuchungen von Flugsaurierproben einen unbestreitbaren Beweis erbracht: Flugsaurier hatten Pyknofasermäntel. Sordes pilosus (was übersetzt “haariger Dämon” bedeutet) und Jeholopterus ninchengensis zeigen Pyknofasern an Kopf und Körper.

Das Vorhandensein von Pyknofasern weist stark darauf hin, dass Flugsaurier endotherm (warmblütig) waren. Sie unterstützten die Wärmeregulierung, wie es bei warmblütigen Tieren üblich ist, die eine Isolierung benötigen, um einen übermäßigen Wärmeverlust zu verhindern. Pyknofasern waren flexible, kurze Filamente, etwa fünf bis sieben Millimeter lang und von relativ einfacher Struktur mit einem hohlen zentralen Kanal. Flugsaurierfelle könnten in ihrer Dichte mit vielen mesozoischen Säugetieren vergleichbar gewesen sein.[b]
Beziehung zu Federn[edit]
Flugsaurierfilamente könnten einen gemeinsamen Ursprung mit Federn haben, wie 2002 von Czerkas und Ji spekuliert.[30] Im Jahr 2009 kam Kellner zu dem Schluss, dass Pyknofasern ähnlich wie die Protofedern der Theropoden aufgebaut sind.[46] Andere waren nicht überzeugt, da der Unterschied zu den “Federkielen” vieler vogelähnlicher Maniraptoran-Exemplare zu grundlegend war.
Eine Studie von 2018 über die Überreste von zwei kleinen Pterosauriern aus dem Jurazeitalter aus der Inneren Mongolei, China, ergab, dass Pterosaurier eine breite Palette von Pyknofaserformen und -strukturen aufwiesen, im Gegensatz zu den homogenen Strukturen, von denen allgemein angenommen wurde, dass sie sie bedecken. Einige von diesen hatten ausgefranste Enden, die in ihrer Struktur vier verschiedenen Federtypen sehr ähnlich waren, die von Vögeln oder anderen Dinosauriern bekannt waren, aber vor der Studie fast nie von Flugsauriern bekannt waren, was auf Homologie schließen lässt.[77][78] Eine Antwort auf diese Studie wurde im Jahr 2020 veröffentlicht, in der vermutet wurde, dass die auf den Anurognathiden beobachteten Strukturen tatsächlich auf die Zersetzung von Aktinofibrillen zurückzuführen sind: eine Art Faser, die zur Verstärkung und Versteifung des Flügels verwendet wird.[79] In einer Antwort darauf weisen die Autoren des Papiers von 2018 jedoch darauf hin, dass sich das Vorhandensein der Strukturen über das Patagium hinaus erstreckt und sowohl Aktinofibrillen als auch Filamente vorhanden sind Jeholopterus ningchengensis[80] und Sordes pilosus.[81] Die verschiedenen Formen der Filamentstruktur, die in der Studie von 2018 auf den Anurognathiden vorhanden waren, würden auch eine Form der Zersetzung erfordern, die die verschiedenen gesehenen “Filamentformen” verursachen würde. Sie kommen daher zu dem Schluss, dass die sparsamste Interpretation der Strukturen darin besteht, dass es sich um filamentöse Protofedern handelt.[82]
Entdeckungsgeschichte[edit]
Erste Funde[edit]

Flugsaurierfossilien sind aufgrund ihrer leichten Knochenkonstruktion sehr selten. Komplette Skelette finden sich in der Regel nur in geologischen Schichten mit außergewöhnlichen Erhaltungsbedingungen, den sogenannten Lagerstätten. Die Stücke von einem solchen LagerstätteDer spätjurassische Solnhofener Kalkstein in Bayern wurde von reichen Sammlern sehr gesucht. Der italienische Naturforscher Cosimo Alessandro Collini war 1784 der erste Wissenschaftler in der Geschichte, der ein Flugsaurierfossil beschrieb. Zu dieser Zeit wurden die Konzepte der Evolution und des Aussterbens nur unvollkommen entwickelt. Der bizarre Körperbau des Flugsauriers war daher schockierend, da er keiner vorhandenen Tiergruppe eindeutig zugeordnet werden konnte. Die Entdeckung von Flugsauriern würde daher eine wichtige Rolle für den Fortschritt der modernen Paläontologie und Geologie spielen. Wenn solche Kreaturen noch lebten, war nur das Meer ein glaubwürdiger Lebensraum, und Collini schlug vor, dass es sich um ein schwimmendes Tier handeln könnte, das seine langen Vorderbeine als Paddel benutzte.[88] Einige Wissenschaftler unterstützten die aquatische Interpretation bis 1830, als der deutsche Zoologe Johann Georg Wagler dies vorschlug Pterodactylus benutzte seine Flügel als Flossen und war mit Ichthyosauria und Plesiosauria verbunden.[89]

Im Jahr 1800 schlug Johann Hermann in einem Brief an Georges Cuvier erstmals vor, dass es sich um eine fliegende Kreatur handelt. Cuvier stimmte 1801 zu und verstand, dass es sich um ein ausgestorbenes fliegendes Reptil handelte.[90] 1809 prägte er den Namen Ptéro-Dactyle, “Flügelfinger”.[91] Dies wurde 1815 latinisiert Pterodactylus.[92] Zuerst wurden die meisten Arten dieser Gattung zugeordnet und schließlich wurde “Pterodaktylus” populär und falsch auf alle Mitglieder von Pterosauria angewendet.[14] Heute beschränken Paläontologen den Begriff auf die Gattung Pterodactylus oder Mitglieder der Pterodactyloidea.[15]
In den Jahren 1812 und 1817 beschrieb Samuel Thomas von Soemmerring das Originalexemplar und ein weiteres neu.[93] Er sah sie als mit Vögeln und Fledermäusen verbunden an. Obwohl er sich darin irrte, würde sein “Fledermausmodell” im 19. Jahrhundert sehr einflussreich sein. 1843 glaubte Edward Newman, Flugsaurier würden Beuteltiere fliegen.[95] Da das Fledermausmodell Flugsaurier korrekt als pelzig und warmblütig darstellte, näherte es sich besser der wahren Physiologie der Flugsaurier als Cuviers “Reptilienmodell”. Johann Jakob Kaup prägte 1834 den Begriff Pterosauria.[96]
Forschung erweitern[edit]

Im Jahr 1828 fand Mary Anning in England die erste Flugsauriergattung außerhalb Deutschlands, die von Richard Owen als benannt wurde Dimorphodon, auch der erste bekannte nicht-pterodaktyloide Flugsaurier. Später im Jahrhundert produzierte das frühe Kreide-Cambridge-Grünsand Tausende von Flugsaurier-Fossilien, die jedoch von schlechter Qualität waren und hauptsächlich aus stark erodierten Fragmenten bestanden. Basierend auf diesen würden dennoch zahlreiche Gattungen und Arten benannt. Viele wurden von Harry Govier Seeley beschrieben, dem damaligen englischen Hauptexperten, der auch das erste Flugsaurierbuch schrieb. Ornithosauria,[100] und 1901 das erste populäre Buch,Drachen der Luft. Seeley hielt Flugsaurier für warmblütige und dynamische Wesen, die eng mit Vögeln verwandt waren.[101] Zuvor hatte der Evolutionist St. George Jackson Mivart vorgeschlagen, Flugsaurier seien die direkten Vorfahren der Vögel.[102] Owen widersetzte sich den Ansichten beider Männer und sah Flugsaurier als kaltblütige “wahre” Reptilien.
In den USA entdeckte Othniel Charles Marsh 1870 Pteranodon in der Niobrara-Kreide, damals der größte bekannte Flugsaurier, der erste zahnlose und der erste aus Amerika. Auch diese Schichten lieferten Tausende von Fossilien, darunter auch relativ vollständige Skelette, die dreidimensional erhalten blieben, anstatt wie bei den Solnhofener Exemplaren stark komprimiert zu werden. Dies führte zu einem viel besseren Verständnis vieler anatomischer Details, wie beispielsweise der hohlen Natur der Knochen.
In der Zwischenzeit wurden Funde aus den Solnhofen fortgesetzt, die den größten Teil der vollständig entdeckten hochwertigen Exemplare ausmachten. Sie erlaubten es, die meisten neuen Basaltaxa zu identifizieren, wie z Rhamphorhynchus, Scaphognathus und Dorygnathus. Dieses Material brachte eine deutsche Schule für Flugsaurierforschung hervor, die fliegende Reptilien als warmblütige, pelzige und aktive mesozoische Gegenstücke moderner Fledermäuse und Vögel ansah. 1882 veröffentlichten Marsh und Karl Alfred Zittel Studien über die Flügelmembranen von Exemplaren von Rhamphorhynchus.[107][108] Die deutschen Studien wurden bis in die 1930er Jahre fortgesetzt und beschreiben neue Arten wie Anurognathus. Im Jahr 1927 entdeckte Ferdinand Broili Haarfollikel in Pterosaurierhaut,[109] und der Paläoneurologe Tilly Edinger stellte fest, dass das Gehirn von Flugsauriern eher dem von Vögeln ähnelte als modernen kaltblütigen Reptilien.[110]
Im Gegensatz dazu verloren englische und amerikanische Paläontologen Mitte des 20. Jahrhunderts weitgehend das Interesse an Flugsauriern. Sie sahen sie als gescheiterte Evolutionsexperimente, kaltblütig und schuppig, die kaum fliegen konnten. Die größeren Arten konnten nur gleiten, mussten auf Bäume klettern oder sich von Klippen werfen, um einen Start zu erreichen. 1914 wurde die Aerodynamik von Flugsauriern erstmals von Ernest Hanbury Hankin und David Meredith Seares Watson quantitativ analysiert, aber interpretiert Pteranodon als reines Segelflugzeug.[111] In den 1940er und 1950er Jahren wurde wenig über die Gruppe geforscht.
Pterosaurier Renaissance[edit]

Die Situation für Dinosaurier war vergleichbar. Ab den 1960er Jahren fand eine Dinosaurier-Renaissance statt, bei der die Anzahl der Studien und kritischen Ideen rasch zunahm, beeinflusst durch die Entdeckung zusätzlicher Fossilien von Deinonychus, dessen spektakuläre Züge die fest verwurzelte Orthodoxie widerlegten. 1970 ebenfalls die Beschreibung des pelzigen Flugsauriers Sordes begann, was Robert Bakker eine Renaissance der Flugsaurier nannte.[112] Insbesondere Kevin Padian propagierte die neuen Ansichten und veröffentlichte eine Reihe von Studien, in denen Flugsaurier als warmblütige, aktive und rennende Tiere dargestellt wurden.[113][114][115] Dies fiel mit einer Wiederbelebung der deutschen Schule durch die Arbeit von Peter Wellnhofer zusammen, der in den 1970er Jahren den Grundstein für die moderne Flugsaurierwissenschaft legte. 1978 veröffentlichte er das erste Flugsaurier-Lehrbuch, das Handbuch der Paläoherptologie, Teil 19: Pterosauria,[117] und 1991 das zweite populärwissenschaftliche Flugsaurierbuch, das Enzyklopädie der Flugsaurier.

Diese Entwicklung beschleunigte sich durch die Nutzung von zwei neuen Lagerstätten. In den 1970er Jahren begann die frühe kreidezeitliche Santana-Formation in Brasilien Kreideknollen zu produzieren, die, obwohl sie oft in ihrer Größe und Vollständigkeit der darin enthaltenen Fossilien begrenzt waren, perfekt erhaltene dreidimensionale Pterosaurier-Skelettteile enthielten. Deutsche und niederländische Institute kauften solche Knötchen von fossilen Wilderern und bereiteten sie in Europa vor, damit ihre Wissenschaftler viele neue Arten beschreiben und eine ganz neue Fauna entdecken konnten. Bald fingen brasilianische Forscher, darunter Alexander Kellner, den Handel ab und nannten noch mehr Arten.
Noch produktiver war die frühe Kreidezeit des chinesischen Jehol Biota von Liaoning, die seit den 1990er Jahren Hunderte von exquisit erhaltenen zweidimensionalen Fossilien hervorgebracht hat, die häufig Weichteilreste aufweisen. Chinesische Forscher wie Lü Junchang haben erneut viele neue Taxa benannt. Als die Entdeckungen auch in anderen Teilen der Welt zunahmen, kam es zu einem plötzlichen Anstieg der Gesamtzahl der genannten Gattungen. Bis 2009, als sie auf etwa neunzig gestiegen waren, zeigte dieses Wachstum keine Anzeichen einer Abflachung.[119] Im Jahr 2013 gab MP Witton an, dass die Zahl der entdeckten Flugsaurierarten auf 130 gestiegen war.[120] Über neunzig Prozent der bekannten Taxa wurden während der “Renaissance” benannt. Viele von ihnen stammten aus Gruppen, deren Existenz unbekannt war. Fortschritte in der Rechenleistung ermöglichten es, ihre komplexen Beziehungen durch die quantitative Methode der Kladistik zu bestimmen. Neue und alte Fossilien lieferten viel mehr Informationen, wenn sie moderner UV-Fotografie oder Röntgenfotografie oder CAT-Scans unterzogen wurden. Erkenntnisse aus anderen Bereichen der Biologie wurden auf die erhaltenen Daten angewendet. All dies führte zu erheblichen Fortschritten in der Flugsaurierforschung, wodurch ältere Berichte in populärwissenschaftlichen Büchern völlig veraltet waren.
Evolution und Aussterben[edit]
Ursprünge[edit]

Da die Anatomie der Flugsaurier für den Flug so stark verändert wurde und unmittelbare fossile Übergangsvorgänger bisher nicht beschrieben wurden, ist die Abstammung der Flugsaurier nicht vollständig verstanden. Die ältesten bekannten Flugsaurier waren bereits vollständig an einen fliegenden Lebensstil angepasst. Seit Seeley wurde erkannt, dass Flugsaurier wahrscheinlich ihren Ursprung in den “Archosauriern” hatten, die heute als Archosauromorpha bezeichnet werden. In den 1980er Jahren ergaben frühe kladistische Analysen, dass es sich um Avemetatarsalier handelte (Archosaurier, die näher an Dinosauriern als an Krokodilen waren). Da dies sie auch zu ziemlich nahen Verwandten der Dinosaurier machen würde, wurden diese Ergebnisse von Kevin Padian als Bestätigung seiner Interpretation von Flugsauriern als zweibeinige warmblütige Tiere angesehen. Da diese frühen Analysen auf einer begrenzten Anzahl von Taxa und Zeichen basierten, waren ihre Ergebnisse von Natur aus unsicher. Mehrere einflussreiche Forscher, die Padians Schlussfolgerungen ablehnten, boten alternative Hypothesen an. David Unwin schlug eine Abstammung unter den basalen Archosauromorpha vor, insbesondere langhalsige Formen (“Protorosaurier”) wie Tanystropheiden. Eine Platzierung unter basalen Archosauriformen wie Euparkeria wurde auch vorgeschlagen.[22] Einige basale Archosauromorphe scheinen auf den ersten Blick aufgrund ihrer langgliedrigen Anatomie gute Kandidaten für nahe Verwandte von Flugsauriern zu sein. Ein Beispiel ist Sharovipteryx, ein “Protorosaurier” mit Hautmembranen an den Hinterbeinen, die wahrscheinlich zum Gleiten verwendet werden. Eine Studie von Michael Benton aus dem Jahr 1999 ergab, dass Flugsaurier Avemetatarsalier waren, die eng mit ihnen verwandt waren Scleromochlus, und nannte die Gruppe Ornithodira, um Flugsaurier und Dinosaurier zu umfassen.[124]

Zwei Forscher, S. Christopher Bennett im Jahr 1996,[125] und der Paläo-Künstler David Peters veröffentlichten im Jahr 2000 Analysen, in denen festgestellt wurde, dass Flugsaurier Protorosaurier sind oder eng mit ihnen verwandt sind. Peters sammelte jedoch neuartige anatomische Daten mithilfe einer nicht verifizierten Technik namens “Digital Graphic Segregation” (DGS), bei der Bilder von Pterosaurier-Fossilien mithilfe einer Fotobearbeitungssoftware digital verfolgt werden.[126] Bennett erholte Flugsaurier erst als nahe Verwandte der Protorosaurier, nachdem er die Merkmale des Hinterbeins aus seiner Analyse entfernt hatte, um die Möglichkeit einer auf Fortbewegung basierenden konvergenten Evolution zwischen Flugsauriern und Dinosauriern zu testen. Eine Antwort von Dave Hone und Michael Benton aus dem Jahr 2007 konnte dieses Ergebnis nicht reproduzieren und stellte fest, dass Flugsaurier auch ohne Hinterbeincharakter eng mit Dinosauriern verwandt sind. Sie kritisierten auch David Peters dafür, dass er Schlussfolgerungen ohne Zugang zu den primären Beweisen, dh den Pterosaurier-Fossilien selbst, gezogen hatte.[127] Hone und Benton kamen zu dem Schluss, dass, obwohl mehr basale Pterosauromorphe erforderlich sind, um ihre Beziehungen zu klären, aktuelle Erkenntnisse darauf hinweisen, dass Pterosaurier Avemetatarsalier sind, wie entweder die Schwestergruppe von Scleromochlus oder ein Zweig zwischen letzterem und Lagosuchus.[127] Eine archosaurierorientierte phylogenetische Analyse von Sterling Nesbitt aus dem Jahr 2011 profitierte von weitaus mehr Daten und fand jedoch starke Unterstützung für Flugsaurier als Avemetatarsalier Scleromochlus wurde wegen seiner schlechten Konservierung nicht berücksichtigt.[128] Eine archosauromorph-fokussierte Studie von Martin Ezcurra aus dem Jahr 2016 umfasste verschiedene vorgeschlagene Pterosaurier-Verwandte, fand jedoch auch heraus, dass Pterosaurier näher an Dinosauriern sind und nichts mit basaleren Taxa zu tun haben.[129] Ausgehend von seiner Analyse von 1996 veröffentlichte Bennett eine Studie zu 2020 über Scleromochlus was argumentierte, dass beide Scleromochlus und Flugsaurier waren Nicht-Archosaurier-Archosauromorphe, wenn auch nicht besonders eng miteinander verwandt.[130]
Ein verwandtes Problem ist der Ursprung des Flugsaurierfluges. Wie bei Vögeln können Hypothesen in zwei Hauptsorten eingeteilt werden: “gemahlen” oder “Baum runter”. Das Klettern auf einen Baum würde dazu führen, dass Höhe und Schwerkraft sowohl die Energie als auch einen starken Selektionsdruck für den beginnenden Flug liefern. Rupert Wild schlug 1983 einen hypothetischen “Propterosaurus” vor: ein eidechsenartiges Baumtier, das eine Membran zwischen seinen Gliedmaßen entwickelt, um zuerst sicher zu springen und dann den vierten Finger allmählich zu verlängern, um zu gleiten.[132] Nachfolgende kladistische Ergebnisse passten jedoch nicht gut zu diesem Modell. Weder Protorosaurier noch Ornithodirane sind Eidechsen biologisch gleichwertig. Darüber hinaus ist der Übergang zwischen Gleit- und Schlagflug nicht gut verstanden. Neuere Studien zur Morphologie der basalen Pterosaurier-Hinterbeine scheinen eine Verbindung zu bestätigen Scleromochlus. Wie dieser Archosaurier haben basale Flugsaurierlinien plantigrade Hinterbeine, die Anpassungen für die Salzbildung zeigen.[133]
Aussterben[edit]

Es wurde einmal angenommen, dass die Konkurrenz mit Frühaufsteherarten zum Aussterben vieler Flugsaurier geführt haben könnte.[134] Ein Teil davon ist auf die Tatsache zurückzuführen, dass früher angenommen wurde, dass bis zum Ende der Kreidezeit nur noch große Arten von Flugsauriern vorhanden waren (nicht mehr wahr; siehe unten). Es wurde angenommen, dass die kleineren Arten ausgestorben sind und ihre Nische von Vögeln gefüllt ist.[135] Der Rückgang der Flugsaurier (falls tatsächlich vorhanden) scheint jedoch nichts mit der Vogelvielfalt zu tun zu haben, da die ökologische Überschneidung zwischen den beiden Gruppen minimal zu sein scheint.[136] Tatsächlich wurden zumindest einige Vogelnischen vor dem KT-Ereignis von Flugsauriern zurückerobert.[137] Am Ende der Kreidezeit scheint das Aussterben der Kreidezeit und des Paläogens, bei dem alle Nicht-Vogel-Dinosaurier und die meisten Vogel-Dinosaurier sowie viele andere Tiere ausgelöscht wurden, auch die Flugsaurier gefangen zu haben.
In den frühen 2010er Jahren wurden mehrere neue Flugsaurier-Taxa entdeckt, die aus dem Campanian / Maastrichtian stammen, wie beispielsweise die Ornithocheiriden Piksi und “Ornithocheirus“, mögliche Pteranodontiden und Nyctosauriden, mehrere Tapejariden und das unbestimmte Nicht-Azhdarchid Navajodactylus.[138][139] Kleine azhdarchoide Flugsaurier waren auch im Campanian vorhanden. Dies deutet darauf hin, dass die Pterosaurierfaunen der späten Kreidezeit weitaus vielfältiger waren als bisher angenommen und möglicherweise nicht einmal signifikant gegenüber der frühen Kreidezeit zurückgegangen sind.
In der Csehbánya-Formation waren offenbar kleine Flugsaurierarten vorhanden, was auf eine höhere Vielfalt von Flugsauriern aus der späten Kreidezeit hinweist als bisher.[140] Die jüngsten Ergebnisse eines kleinen erwachsenen Azhdarchiden in Katzengröße deuten ferner darauf hin, dass kleine Flugsaurier aus der späten Kreidezeit im Fossilienbestand möglicherweise nur selten erhalten geblieben sind, was durch die Tatsache unterstützt wird, dass eine starke Tendenz gegen terrestrische kleine Wirbeltiere wie Jungtiere besteht Dinosaurier, und dass ihre Vielfalt tatsächlich viel größer gewesen sein könnte als bisher angenommen.[141]
Zumindest einige nicht-pterodaktyloide Flugsaurier überlebten bis in die späte Kreidezeit und postulierten eine Lazarus-Taxa-Situation für spätkreidezeitliche Flugsaurierfaunen.[142]
Klassifikation und Phylogenie[edit]
In der phylogenetischen Taxonomie wurde die Klade Pterosauria normalerweise als knotenbasiert definiert und in mehreren ausführlich untersuchten Taxa sowie in Taxa verankert, die als primitiv gelten. Eine Studie aus dem Jahr 2003 definierte Pterosauria als “den jüngsten gemeinsamen Vorfahren der Anurognathidae, Preondactylus und Quetzalcoatlus und alle ihre Nachkommen. “[143] Diese Arten der Definition würden jedoch zwangsläufig alle verwandten Arten, die etwas primitiver sind, aus der Pterosauria herauslassen. Um dies zu beheben, wurde eine neue Definition vorgeschlagen, die den Namen nicht an eine bestimmte Art, sondern an ein anatomisches Merkmal, das Vorhandensein eines vergrößerten vierten Fingers, der eine Flügelmembran trägt, verankert.[144] Diese “Apomorophy-basierte” Definition wurde 2020 vom PhyloCode übernommen.[145] Eine breitere Gruppe, Pterosauromorphawurde definiert als alle Ornithodirane, die enger mit Flugsauriern verwandt sind als mit Dinosauriern.[146]
Die interne Klassifizierung von Flugsauriern war historisch schwierig, da es viele Lücken im Fossilienbestand gab. Ab dem 21. Jahrhundert füllen neue Entdeckungen diese Lücken und geben ein besseres Bild der Entwicklung der Flugsaurier. Traditionell waren sie in zwei Unterordnungen unterteilt: die Rhamphorhynchoidea, eine “primitive” Gruppe von Langschwanz-Flugsauriern, und die Pterodactyloidea, “fortgeschrittene” Flugsaurier mit kurzen Schwänzen.[22] Diese traditionelle Aufteilung wurde jedoch weitgehend aufgegeben. Rhamphorhynchoidea ist eine paraphyletische (unnatürliche) Gruppe, da sich die Pterodactyloide direkt aus ihnen und nicht aus einem gemeinsamen Vorfahren entwickelt haben. Mit zunehmendem Einsatz von Kladistiken ist sie bei den meisten Wissenschaftlern in Ungnade gefallen.[120][147]
Die genauen Beziehungen zwischen Flugsauriern sind noch ungeklärt. Viele Studien zu Pterosaurier-Beziehungen in der Vergangenheit enthielten nur begrenzte Daten und waren sehr widersprüchlich. Neuere Studien mit größeren Datenmengen beginnen jedoch, die Dinge klarer zu machen. Das folgende Cladogramm (Stammbaum) folgt einer phylogenetischen Analyse, die 2018 von Longrich, Martill und Andres vorgelegt wurde.[137]
Paläobiologie[edit]


Flug[edit]
Die Mechanik des Flugsaurierfluges ist derzeit noch nicht vollständig verstanden oder modelliert.[148][149]
Katsufumi Sato, ein japanischer Wissenschaftler, führte Berechnungen mit modernen Vögeln durch und kam zu dem Schluss, dass es für einen Flugsaurier unmöglich war, in der Luft zu bleiben.[148] Im Buch Haltung, Fortbewegung und Paläoökologie von Flugsauriern Es wird vermutet, dass sie aufgrund der sauerstoffreichen, dichten Atmosphäre der späten Kreidezeit fliegen konnten.[150] Allerdings haben sowohl Sato als auch die Autoren von Haltung, Fortbewegung und Paläoökologie von Flugsauriern Ihre Forschung stützte sich auf die inzwischen veralteten Theorien, dass Flugsaurier wie Seevögel aussehen, und die Größenbeschränkung gilt nicht für terrestrische Flugsaurier wie Azhdarchiden und Tapejariden. Darüber hinaus kam Darren Naish zu dem Schluss, dass atmosphärische Unterschiede zwischen der Gegenwart und dem Mesozoikum für die riesige Größe der Flugsaurier nicht erforderlich sind.[151]

Ein weiteres Problem, das schwer zu verstehen war, ist, wie sie gestartet sind. Frühere Vorschläge waren, dass Flugsaurier größtenteils kaltblütige gleitende Tiere waren, die wie moderne Eidechsen Wärme aus der Umgebung bezogen, anstatt Kalorien zu verbrennen. In diesem Fall war unklar, wie die größeren von enormer Größe mit einem ineffizienten kaltblütigen Stoffwechsel eine vogelähnliche Startstrategie verwalten konnten, bei der nur die Hinterbeine verwendet wurden, um Schub zu erzeugen, um in die Luft zu gelangen. Spätere Untersuchungen zeigen, dass sie stattdessen warmblütig sind, starke Flugmuskeln haben und die Flugmuskeln zum Gehen als Vierbeiner verwenden.[152]Mark Witton von der University of Portsmouth und Mike Habib von der Johns Hopkins University schlugen vor, dass Flugsaurier einen Voltigiermechanismus verwenden, um einen Flug zu erhalten.[153] Die enorme Kraft ihrer geflügelten Vorderbeine würde es ihnen ermöglichen, mit Leichtigkeit abzuheben.[152] In der Luft könnten Flugsaurier Geschwindigkeiten von bis zu 120 km / h erreichen und Tausende von Kilometern zurücklegen.[153]
1985 beauftragte die Smithsonian Institution den Luftfahrtingenieur Paul MacCready mit dem Bau eines Arbeitsmodells von Quetzalcoatlus northropi. Die Replik wurde mit einer bodengestützten Winde gestartet. Es flog 1986 mehrmals und wurde im Rahmen des IMAX-Films von Smithsonian gedreht Auf dem Flügel.[154][155]
Luftsäcke und Atmung[edit]
Eine Studie aus dem Jahr 2009 zeigte, dass Flugsaurier ein Lungen-Luft-Sack-System und eine präzise gesteuerte Skelettatmungspumpe hatten, die ein Durchfluss-Lungenbeatmungsmodell bei Flugsauriern unterstützt, das dem von Vögeln entspricht. Die Anwesenheit von a subkutan Das Luftsacksystem in mindestens einigen Pterodactyloiden hätte die Dichte des lebenden Tieres weiter verringert.[47] Wie moderne Krokodile schienen Flugsaurier einen Leberkolben zu haben, da ihre Schulter-Brust-Gürtel zu unflexibel waren, um das Brustbein wie bei Vögeln zu bewegen, und sie besaßen starke Gastralien.[156] Somit hatte ihr Atmungssystem Eigenschaften, die mit beiden modernen Archosaurierkladen vergleichbar waren.
Nervöses System[edit]
Eine Röntgenuntersuchung der Gehirnhöhlen von Flugsauriern ergab, dass die Tiere (Rhamphorhynchus muensteri und Anhanguera santanae) hatten massive Flocken. Der Flockulus ist eine Gehirnregion, die Signale von Gelenken, Muskeln, Haut und Gleichgewichtsorganen integriert.[16] Die Flockchen der Flugsaurier nahmen 7,5% der gesamten Gehirnmasse der Tiere ein, mehr als bei jedem anderen Wirbeltier. Vögel haben im Vergleich zu anderen Tieren ungewöhnlich große Flocken, die jedoch nur zwischen 1 und 2% der gesamten Gehirnmasse ausmachen.[16]
Der Flockulus sendet neuronale Signale aus, die kleine automatische Bewegungen in den Augenmuskeln erzeugen. Diese halten das Bild auf der Netzhaut eines Tieres stabil. Flugsaurier hatten möglicherweise aufgrund ihrer großen Flügelgröße einen so großen Flockulus, was bedeuten würde, dass viel mehr sensorische Informationen verarbeitet werden müssten.[16] Die geringe relative Masse der Flocken bei Vögeln ist auch auf Vögel zurückzuführen, die insgesamt ein viel größeres Gehirn haben. Obwohl dies als Hinweis darauf angesehen wurde, dass Flugsaurier in einer strukturell einfacheren Umgebung lebten oder im Vergleich zu Vögeln ein weniger komplexes Verhalten zeigten,[157] Jüngste Studien an Krokodilen und anderen Reptilien zeigen, dass Sauropsiden häufig mit kleinen Gehirnen ein hohes Intelligenzniveau erreichen.[158] Studien zum Endocast von Allkaruen zeigen, dass die Gehirnentwicklung bei Pterodaktyloiden ein modularer Prozess war.[159]
Bodenbewegung[edit]

Die Hüftpfannen der Flugsaurier sind leicht nach oben gerichtet, und der Kopf des Femurs (Oberschenkelknochen) ist nur mäßig nach innen gerichtet, was darauf hindeutet, dass die Flugsaurier eine aufrechte Haltung hatten. Es wäre möglich gewesen, den Oberschenkel während des Fluges in eine horizontale Position zu heben, wie es gleitende Eidechsen tun.
Es gab erhebliche Debatten darüber, ob Flugsaurier als Vierbeiner oder als Zweibeiner unterwegs waren. In den 1980er Jahren schlug der Paläontologe Kevin Padian kleinere Flugsaurier mit längeren Hinterbeinen vor, wie z Dimorphodon, könnte zusätzlich zum Fliegen wie Straßenläufer gelaufen oder sogar zweibeinig gelaufen sein.[115] Später wurde jedoch eine große Anzahl von Flugsaurierbahnen mit einem charakteristischen Vierzehen-Hinterfuß und einem Dreizehen-Vorderfuß gefunden. Dies sind die unverkennbaren Drucke von Flugsauriern, die auf allen Vieren gehen.[160][161]
Fossile Fußabdrücke zeigen, dass Flugsaurier mit dem gesamten Fuß in Bodenkontakt standen (Plantigrade), ähnlich wie bei vielen Säugetieren wie Menschen und Bären. Fußabdrücke von Azhdarchiden und mehreren nicht identifizierten Arten zeigen, dass Flugsaurier mit einer aufrechten Haltung gingen, wobei ihre vier Gliedmaßen fast senkrecht unter dem Körper gehalten wurden. Dies ist eine energieeffiziente Haltung, die von den meisten modernen Vögeln und Säugetieren anstelle der ausgestreckten Gliedmaßen moderner Reptilien verwendet wird.[69][152] In der Tat können aufrechte Gliedmaßen bei Flugsauriern allgegenwärtig sein.[133]

Obwohl die Anatomie einiger Flugsaurier (insbesondere Pterodaktyloide) traditionell als unbeholfen und umständlich am Boden dargestellt wird, deutet dies darauf hin, dass sie kompetente Wanderer und Läufer waren.[162] Frühe Flugsaurier wurden aufgrund des Vorhandenseins einer großen Cruropatagie lange Zeit als besonders umständliche Lokomotiven angesehen, aber auch sie scheinen vor Ort im Allgemeinen effizient gewesen zu sein.[133]
Die Vorderbeinknochen von Azhdarchiden und Ornithocheiriden waren im Vergleich zu anderen Flugsauriern ungewöhnlich lang, und bei Azhdarchiden waren die Knochen von Arm und Hand (Metacarpalen) besonders langgestreckt. Darüber hinaus waren die azhdarchidischen Vorderbeine insgesamt ähnlich wie die schnell laufenden Huftier-Säugetiere proportioniert. Ihre Hinterbeine hingegen waren nicht auf Geschwindigkeit ausgelegt, aber sie waren im Vergleich zu den meisten Flugsauriern lang und ermöglichten eine lange Schrittlänge. Während azhdarchidische Flugsaurier wahrscheinlich nicht rennen konnten, wären sie relativ schnell und energieeffizient gewesen.[69]
Die relative Größe der Hände und Füße bei Flugsauriern (im Vergleich zu modernen Tieren wie Vögeln) kann auf die Art der am Boden geführten Lebensstil-Flugsaurier hinweisen. Azhdarchid Pterosaurier hatten im Vergleich zu ihrer Körpergröße und Beinlänge relativ kleine Füße, wobei die Fußlänge nur etwa 25–30% der Länge des Unterschenkels betrug. Dies deutet darauf hin, dass Azhdarchiden besser für das Gehen auf trockenem, relativ festem Boden geeignet waren. Pteranodon hatten etwas größere Füße (47% der Länge der Tibia), während Pterosaurier mit Filterfütterung wie die Ctenochasmatoide sehr große Füße hatten (69% der Länge der Tibia in Pterodactylus84% in Pterodaustro), angepasst an weiche, schlammige Böden, ähnlich wie moderne Watvögel.[69] Basal-Flugsaurier sind zwar eindeutig auf Vorderbeinen basierende Trägerraketen, haben aber gut zum Hüpfen angepasste Hinterbeine, was auf eine Verbindung mit Archosauriern wie z Scleromochlus.[133]
Schwimmen[edit]
Spuren von Ctenochasmatoiden weisen darauf hin, dass diese Flugsaurier mit ihren Hinterbeinen schwammen. Im Allgemeinen haben diese große Hinterfüße und lange Oberkörper, was darauf hinweist, dass sie wahrscheinlich besser zum Schwimmen geeignet waren als andere Flugsaurier.[163] Pteranodontianer haben umgekehrt mehrere Spezifikationen in ihren Humeri, die als Hinweis auf eine wasserbasierte Version des typischen Vierbeinstarts interpretiert wurden, und einige wie Boreopteriden müssen beim Schwimmen gefressen haben, da sie nicht in der Lage zu sein scheinen, wie Fregattvögel aus der Luft zu feilschen.[163] Diese Anpassungen sind auch bei terrestrischen Flugsauriern wie Azhdarchiden zu beobachten, die vermutlich noch aus dem Wasser starten mussten, falls sie sich darin befanden. Der Nyctosaurier Alcione kann Anpassungen für das Tauchen mit Flügelantrieb wie moderne Tölpel und Tropikvögel anzeigen.[164]
Diät- und Ernährungsgewohnheiten[edit]
Traditionell wurden fast alle Flugsaurier als oberflächenfressende Fischfresser oder Fischfresser angesehen, eine Ansicht, die immer noch die Populärwissenschaft dominiert. Heutzutage wird angenommen, dass viele Flugsauriergruppen terrestrische Fleischfresser, Allesfresser oder Insektenfresser waren.
Schon früh wurde erkannt, dass die kleinen Anurognathidae nachtaktive Insektenfresser aus der Luft waren. Mit hochflexiblen Gelenken am Flügelfinger, einer breiten, dreieckigen Flügelform, großen Augen und einem kurzen Schwanz waren diese Flugsaurier wahrscheinlich analog zu Nachtgläsern oder vorhandenen insektenfressenden Fledermäusen und konnten bei relativ niedrigen Geschwindigkeiten eine hohe Manövrierfähigkeit erreichen.[165]
Die Interpretation der Gewohnheiten von Basalgruppen hat sich grundlegend geändert. Dimorphodon, in der Vergangenheit als Papageientaucher-Analogon gedacht, wird durch seine Kieferstruktur, seinen Gang und seine schlechten Flugfähigkeiten als terrestrisches / semiarboreales Raubtier kleiner Säugetiere, Squamate und großer Insekten angezeigt. Sein robustes Gebiss verursacht Campylognathoides Als Generalist oder terrestrisches Raubtier kleiner Wirbeltiere gesehen zu werden, aber der äußerst robuste Humerus und die hochgradige Flügelmorphologie lassen darauf schließen, dass es in der Lage gewesen sein könnte, Beute auf dem Flügel zu fangen. Der kleine Insektenfresser Carniadactylus und je größer Eudimorphodon waren hochfliegende Tiere und schnelle, bewegliche Flieger mit langen, robusten Flügeln. Eudimorphodon wurde mit Fischresten im Magen gefunden, aber sein Gebiss deutet auf eine opportunistische Ernährung hin. Schlankflügelig Austriadactylus und Caviramus waren wahrscheinlich terrestrische / semiarboreale Generalisten. Caviramus hatte wahrscheinlich eine starke Beißkraft, was auf eine Anpassung an harte Lebensmittel hinweist, die angesichts des Zahnverschleißes gekaut worden sein könnten.
Einige Rhamphorhynchidae, wie z Rhamphorhynchus selbst oder Dorygnathuswaren Fischesser mit langen, schlanken Flügeln, nadelartigem Gebiss und langen, dünnen Kiefern. Sericipterus, Scaphognathus und Harpactognathus hatte robustere Kiefer und Zähne (die ziphodont, dolchförmig waren, in Sericipterus) und kürzere, breitere Flügel. Dies waren entweder terrestrische / Luft-Raubtiere von Wirbeltieren[169] oder korvidähnliche Generalisten. Wukongopteridae mögen Darwinopterus wurden zuerst als Raubtiere aus der Luft betrachtet. Ohne robuste Kieferstruktur oder kraftvolle fliegende Muskeln gelten sie heute als baumartige oder semiterrestrische Insektenfresser. Darwinopterus robustidensinsbesondere scheint ein Käferspezialist gewesen zu sein.[171]
Bei Pterodactyloiden ist eine größere Variation in der Ernährung vorhanden. Pteranodontia enthielt viele fischfressende Taxa, wie die Ornithocheirae, Boreopteridae, Pteranodontidae und Nyctosauridae. Aufgrund der Nischenaufteilung waren Ornithocheire und die späteren Nyctosauriden Luftdip-Feeder wie die heutigen Fregattvögel (mit Ausnahme des angepassten Tauchens) Alcione elainus), während Boreopteriden Süßwassertauchertiere waren, die Kormoranen ähnlich waren, und pelagische Taucher der Pteranodonten, die Tölpeln und Tölpeln ähnelten. Die Istiodactylide waren wahrscheinlich hauptsächlich Aasfresser. Archaeopterodactyloidea erhielt Nahrung in Küsten- oder Süßwasserlebensräumen. Germanodactylus und Pterodactylus waren Fischfresser, während die Ctenochasmatidae Suspensionsfresser waren und ihre zahlreichen feinen Zähne verwendeten, um kleine Organismen aus seichtem Wasser zu filtern. Pterodaustro wurde für die flamingoähnliche Filterfütterung angepasst.
Im Gegensatz dazu waren Azhdarchoidea meist terrestrische Flugsaurier. Tapejaridae waren baumartige Allesfresser, die Samen und Früchte mit kleinen Insekten und Wirbeltieren ergänzten.[163][174]Dsungaripteridae waren spezialisierte Weichtierfresser, die mit ihren kräftigen Kiefern die Schalen von Weichtieren und Krebstieren zerdrückten. Thalassodromidae waren wahrscheinlich terrestrische Fleischfresser. Thalassodromeus selbst wurde nach einer als “Skim-Fütterung” bekannten Fangmethode benannt, die später als biomechanisch unmöglich angesehen wurde. Vielleicht verfolgte es angesichts seiner verstärkten Kiefergelenke und seiner relativ hohen Beißkraft eine relativ große Beute.[175]Azhdarchidae werden heute als terrestrische Raubtiere verstanden, die gemahlenen Nashornvögeln oder einigen Störchen ähneln und alle Beutetiere fressen, die sie als Ganzes schlucken könnten.[176]Hatzegopteryx war ein robust gebautes Raubtier relativ großer Beute, einschließlich mittelgroßer Dinosaurier.[177][178]Alanqa Möglicherweise war er ein spezialisierter Weichtierfresser.[179] Eine erstaunliche Studie wurde in der Zeitschrift Cretaceous Research veröffentlicht. Paläontologen analysierten den Kiefer einer Flugsaurierart Lonchodraco giganteus und fanden Foramina nahe der Schnabelspitze. Sie zeigten ein ähnliches Muster wie vorhandene Vögel, was auf eine taktile Fütterungsgewohnheit bei Flugsauriern hindeuten würde.[180]
Natürliche Raubtiere[edit]
Es ist bekannt, dass Flugsaurier von Theropoden gefressen wurden. In der Ausgabe vom 1. Juli 2004 von NaturDer Paläontologe Eric Buffetaut diskutiert höchstwahrscheinlich ein Fossil aus der frühen Kreidezeit von drei Halswirbeln eines Flugsauriers mit dem gebrochenen Zahn eines Spinosauriers Irritator, eingebettet darin. Es ist bekannt, dass die Wirbel nicht gefressen und der Verdauung ausgesetzt wurden, da die Gelenke immer noch artikuliert sind.[181]
Fortpflanzung und Lebensgeschichte[edit]

Obwohl nur sehr wenig über die Fortpflanzung von Flugsauriern bekannt ist, wird angenommen, dass ähnlich wie bei allen Dinosauriern alle Flugsaurier durch Eiablage reproduziert werden, obwohl solche Befunde sehr selten sind. Das erste bekannte Flugsaurier-Ei wurde in den Steinbrüchen von Liaoning gefunden, wo auch gefiederte Dinosaurier lebten. Das Ei wurde flach gequetscht, ohne Anzeichen von Rissen, so dass die Eier offensichtlich ledrige Schalen hatten, wie bei modernen Eidechsen.[182] Dies wurde durch die Beschreibung eines zusätzlichen Flugsaurier-Eies der Gattung gestützt Darwinopterus, beschrieben im Jahr 2011, das ebenfalls eine ledrige Schale hatte und wie moderne Reptilien, aber im Gegensatz zu Vögeln, im Vergleich zur Größe der Mutter ziemlich klein war.[183] Im Jahr 2014 fünf nicht abgeflachte Eier der Art Hamipterus tianshanensis wurden in einer frühen Kreidelagerstätte im Nordwesten Chinas gefunden. Die Untersuchung der Schalen durch Rasterelektronenmikroskopie zeigte das Vorhandensein einer dünnen kalkhaltigen Eierschalenschicht mit einer Membran darunter.[184] Eine 2007 veröffentlichte Studie über die Struktur und Chemie von Pterosaurier-Eierschalen ergab, dass Pterosaurier wahrscheinlich ihre Eier wie moderne Krokodile und Schildkröten begraben haben. Das Vergraben von Eiern wäre für die frühe Entwicklung von Flugsauriern von Vorteil gewesen, da es mehr gewichtsreduzierende Anpassungen ermöglicht, aber diese Fortpflanzungsmethode hätte auch die Vielfalt der Umgebungen, in denen Flugsaurier leben könnten, eingeschränkt und sie möglicherweise benachteiligt, wenn Sie wurden einer ökologischen Konkurrenz durch Vögel ausgesetzt.[185]
EIN Darwinopterus Das Exemplar zeigt, dass zumindest einige Flugsaurier ein Paar funktioneller Eierstöcke hatten, im Gegensatz zu dem einzelnen funktionellen Eierstock bei Vögeln, was die Reduzierung funktioneller Eierstöcke als Voraussetzung für einen Motorflug abtat.[186]
In Pterosaurier-Embryonen konservierte Flügelmembranen sind gut entwickelt, was darauf hindeutet, dass Pterosaurier kurz nach der Geburt flugbereit waren.[187] Tomographie-Scans von versteinerten Hamipterus Eier deuten darauf hin, dass die jungen Flugsaurier gut entwickelte Oberschenkelknochen zum Gehen hatten, aber schwache Brust zum Fliegen.[188] Es ist nicht bekannt, ob dies für andere Flugsaurier gilt. Es wurden Fossilien von Flugsauriern gefunden, die nur wenige Tage bis eine Woche alt sind (sogenannte “Flaplings”) und mehrere Flugsaurierfamilien repräsentieren, darunter Pterodaktyliden, Rhamphorhinchiden, Ctenochasmatiden und Azhdarchiden.[22] Alle konservieren Knochen, die einen relativ hohen Härtungsgrad aufweisen (Ossifikation) für ihr Alter und Flügelproportionen ähnlich wie bei Erwachsenen. Tatsächlich wurden viele Flugsaurierklappen in der Vergangenheit als Erwachsene betrachtet und in verschiedene Arten eingeteilt. Darüber hinaus finden sich Flatter normalerweise in denselben Sedimenten wie Erwachsene und Jugendliche derselben Art, wie z Pterodactylus und Rhamphorhynchus im Solnhofener Kalkstein Deutschlands gefundene Klappen und Pterodaustro Flaplings aus Argentinien. Alle sind in tiefen Gewässern weit weg von der Küste zu finden.[189]
Für die Mehrheit der Flugsaurierarten ist nicht bekannt, ob sie irgendeine Form der elterlichen Fürsorge praktizierten, aber ihre Fähigkeit zu fliegen, sobald sie aus dem Ei hervorgingen, und die zahlreichen Klappen, die in Umgebungen weit entfernt von Nestern und neben Erwachsenen gefunden wurden, haben die meisten Forscher geführt , einschließlich Christopher Bennett und David Unwin, um zu dem Schluss zu kommen, dass die Jungen für eine relativ kurze Zeit von ihren Eltern abhängig waren, während einer Zeit schnellen Wachstums, während die Flügel lang genug wuchsen, um zu fliegen, und dann das Nest verließen, um für sich selbst zu sorgen , möglicherweise innerhalb von Tagen nach dem Schlüpfen.[22][190] Alternativ können sie in den ersten Lebenstagen gelagerte Eigelbprodukte zur Ernährung verwendet haben, wie dies bei modernen Reptilien der Fall ist, anstatt von den Eltern als Nahrung abhängig zu sein.[189] Versteinert Hamipterus Es wurde gezeigt, dass Nester viele männliche und weibliche Flugsaurier zusammen mit ihren Eiern auf ähnliche Weise wie moderne Seevogelkolonien konservieren.[184][191] Aufgrund der Unterentwicklung der Brust der Jungtiere zum Fliegen wurde dies vorgeschlagen Hamipterus Möglicherweise haben Sie irgendeine Form der elterlichen Fürsorge praktiziert.[188] Diese Studie wurde jedoch inzwischen kritisiert.[192] Die meisten Beweise deuten derzeit darauf hin, dass Flugsaurier-Jungtiere sehr vorsozial sind, ähnlich wie Megapoden-Vögel, die nach dem Schlüpfen ohne elterliche Fürsorge fliegen.
Die Wachstumsraten von Flugsauriern nach dem Schlüpfen waren in verschiedenen Gruppen unterschiedlich. Bei primitiveren Langschwanz-Flugsauriern (“Rhamphorhynchoiden”) wie z RhamphorhynchusDie durchschnittliche Wachstumsrate im ersten Lebensjahr betrug 130% bis 173% und war damit etwas schneller als die Wachstumsrate von Alligatoren. Das Wachstum dieser Arten verlangsamte sich nach der Geschlechtsreife und es hätte mehr als drei Jahre gedauert Rhamphorhynchus maximale Größe erreichen.[190] Im Gegensatz dazu sind die fortgeschritteneren, großen Pterodactyloid-Pterosaurier, wie z Pteranodonwuchs innerhalb des ersten Lebensjahres auf Erwachsenengröße. Zusätzlich hatten Pterodactyloide Wachstum bestimmenDies bedeutet, dass die Tiere eine feste maximale Erwachsenengröße erreichten und aufhörten zu wachsen.[189]
Tägliche Aktivitätsmuster[edit]
Vergleiche zwischen den Skleraringen von Flugsauriern und modernen Vögeln und Reptilien wurden verwendet, um auf die täglichen Aktivitätsmuster von Flugsauriern zu schließen. Die Gattungen der Flugsaurier Pterodactylus, Scaphognathus, und Tupuxuara wurden als tagaktiv gefolgert, Ctenochasma, Pterodaustro, und Rhamphorhynchus wurden als nachtaktiv gefolgert, und Tapejara Es wurde vermutet, dass es kathemeral ist und den ganzen Tag über für kurze Intervalle aktiv ist. Infolgedessen ist das möglicherweise fischfressend Ctenochasma und Rhamphorhynchus Möglicherweise hatten sie ähnliche Aktivitätsmuster wie moderne nachtaktive Seevögel und die Filterfütterung Pterodaustro Möglicherweise hatten sie ähnliche Aktivitätsmuster wie moderne anseriforme Vögel, die nachts fressen. Die Unterschiede zwischen den Aktivitätsmustern der Solnhofener Flugsaurier Ctenochasma, Rhamphorhynchus, Scaphognathus, und Pterodactylus kann auch auf eine Nischenaufteilung zwischen diesen Gattungen hinweisen.[193]
Kulturelle Bedeutung[edit]

Flugsaurier sind seit jeher ein Grundnahrungsmittel der Populärkultur, solange ihre Cousins die Dinosaurier sind, obwohl sie in Filmen, Literatur oder anderen Kunstwerken normalerweise nicht so prominent vertreten sind. Während sich die Darstellung von Dinosauriern in populären Medien als Reaktion auf Fortschritte in der Paläontologie radikal verändert hat, besteht seit Mitte des 20. Jahrhunderts ein hauptsächlich veraltetes Bild von Flugsauriern.[194]
Der vage Oberbegriff “Pterodaktylus” wird häufig für diese Kreaturen verwendet. Die abgebildeten Tiere repräsentieren häufig beides Pteranodon oder (Nicht-Pterodactyloid) Rhamphorhynchusoder eine fiktive Mischung aus beiden.[194] Viele Kinderspielzeuge und Cartoons enthalten “Pterodaktylen” mit Pteranodon-ähnliche Kämme und lang, Rhamphorhynchus– wie Schwänze und Zähne, eine Kombination, die es in der Natur nie gab. Allerdings mindestens ein Flugsaurier tat habe beide die Pteranodon-ähnliche Wappen und Zähne: Ludodactylus, dessen Name wegen seiner Ähnlichkeit mit altem, ungenauem Kinderspielzeug “Spielzeugfinger” bedeutet.[195] Flugsaurier wurden manchmal fälschlicherweise als (die Vorfahren von) Vögeln identifiziert, obwohl Vögel Theropodendinosaurier sind und nicht eng mit Flugsauriern verwandt sind.
Flugsaurier wurden in Sir Arthur Conan Doyles Roman von 1912 in der Fiktion verwendet Die verlorene Welt und seine Verfilmung von 1925. Sie traten seitdem in einer Reihe von Filmen und Fernsehprogrammen auf, darunter im Film von 1933 King Kongund 1966 Eine Million Jahre vor Christus In letzterem Fall musste der Animator Ray Harryhausen seinen Stop-Motion-Modellen ungenaue fledermausähnliche Flügelfinger hinzufügen, um zu verhindern, dass die Membranen auseinanderfallen, obwohl dieser spezielle Fehler in der Kunst bereits vor der Herstellung des Films häufig vorkam. Rodan, ein fiktives Riesenmonster (oder kaiju), die erstmals im Film von 1956 erschien Rodanwird als enorme bestrahlte Art von dargestellt Pteranodon. Rodan ist in mehreren Japanisch erschienen Godzilla Filme, die in den 1960er, 1970er, 1990er und 2000er Jahren veröffentlicht wurden und auch in dem in den USA produzierten Film 2019 erschienen sind Godzilla: König der Monster.[197]
Nach den 1960er Jahren blieben Flugsaurier bis 2001 in bemerkenswerten amerikanischen Filmauftritten größtenteils abwesend Jurassic Park III. Der Paläontologe Dave Hone stellte fest, dass die Flugsaurier in diesem Film nicht wesentlich aktualisiert wurden, um der modernen Forschung Rechnung zu tragen. Anhaltende Fehler waren zahnlose Zähne Pteranodon sollte dargestellt werden, Nistverhalten, von dem bis 2001 bekannt war, dass es ungenau ist, und ledrige Flügel anstelle der straffen Membranen aus Muskelfasern, die für den Flug des Flugsauriers erforderlich sind.[194]
In den meisten Medienauftritten werden Flugsaurier als Fischfresser dargestellt, was nicht ihre volle Ernährungsvariante widerspiegelt. Sie werden auch oft als Raubtiere aus der Luft gezeigt, die Greifvögeln ähneln und menschliche Opfer mit Krallen an den Füßen greifen. Allerdings nur der kleine Anurognathid Vesperopterylus ist dafür bekannt, Greiffüße zu besitzen; Alle anderen Flugsaurier haben flache Plantigradenfüße ohne gegensätzliche Zehen, und die Füße sind im Allgemeinen proportional klein, zumindest im Fall der Pteranodontie.[198]
Siehe auch[edit]
Verweise[edit]
- ^ Andres, B.; Clark, J.; Xu, X. (2014). “Das früheste Pterodactyloid und der Ursprung der Gruppe”. Aktuelle Biologie. 24 (9): 1011–16. doi:10.1016 / j.cub.2014.03.030. PMID 24768054.
- ^ Mark P. Witton (2013), Flugsaurier: Naturgeschichte, Evolution, Anatomie, Princeton University Press, ISBN 978-0-691-15061-1
- ^ David M. Unwin (2010), “Darwinopterus und seine Auswirkungen auf die Pterosaurier-Phylogenie “, Acta Geoscientica Sinica, 31 (1): 68–69
- ^ Jones, Daniel (2003) [1917]Peter Roach; James Hartmann; Jane Setter (Hrsg.), Englisch Aussprachewörterbuch, Cambridge: Cambridge University Press, ISBN 978-3-12-539683-8
- ^ “Flugsaurier”. Merriam-Webster-Wörterbuch.
- ^ “Pterosaurierverteilung in Zeit und Raum: ein Atlas” (PDF). Zitteliana: 61–107. 2008.
- ^ Elgin RA, Hone DW, Frey E (2011). “Das Ausmaß der Flugsaurier-Flugmembran”. Acta Palaeontologica Polonica. 56 (1): 99–111. doi:10.4202 / App.2009.0145.
- ^ “Pterosaur.net :: Terrestrische Fortbewegung”. pterosaur.net. Abgerufen 2020-02-01.
- ^ Geggel 2018-12-17T19: 23: 17Z, Laura. “Es ist offiziell: Diese fliegenden Reptilien, die Pterosaurier genannt werden, waren mit flauschigen Federn bedeckt.”. livingcience.com. Abgerufen 2020-02-01.
- ^ ein b Wang, X.; Kellner, AWA; Zhou, Z.; Campos, DA (2008). “Entdeckung eines seltenen arborealen waldbewohnenden fliegenden Reptils (Pterosauria, Pterodactyloidea) aus China”. Verfahren der Nationalen Akademie der Wissenschaften. 105 (6): 1983–87. Bibcode:2008PNAS..105.1983W. doi:10.1073 / pnas.0707728105. PMC 2538868. PMID 18268340.
- ^ Lawson DA (März 1975). “Flugsaurier aus der neuesten Kreidezeit Westtexas: Entdeckung der größten fliegenden Kreatur”. Wissenschaft. 187 (4180): 947–948. Bibcode:1975Sci … 187..947L. doi:10.1126 / science.187.4180.947. PMID 17745279. S2CID 46396417.
- ^ Buffetaut E, Grigorescu D, Csiki Z (April 2002). “Ein neuer Riesen-Flugsaurier mit einem robusten Schädel aus der jüngsten Kreidezeit Rumäniens”. Naturwissenschaften. 89 (4): 180–84. Bibcode:2002NW ….. 89..180B. doi:10.1007 / s00114-002-0307-1. PMID 12061403. S2CID 15423666.
- ^ Benton, Michael J. (2004). “Ursprung und Beziehungen der Dinosaurier”. In Weishampel, David B.; Dodson, Peter; Osmólska, Halszka (Hrsg.). Die Dinosaurier (2. Aufl.). Berkeley: University of California Press. pp. 7–19. ISBN 978-0-520-24209-8.
- ^ ein b Naisch, Darren. “Flugsaurier: Mythen und Missverständnisse”. Pterosaur.net. Abgerufen 18. Juni 2011.
- ^ ein b Alexander, David E. & Vogel, Steven (2004). Flyer der Natur: Vögel, Insekten und die Biomechanik des Fluges. JHU Drücken Sie. p. 191. ISBN 978-0-8018-8059-9.
- ^ ein b c d Witmer LM, Chatterjee S., Franzosa J., Rowe T. (2003). “Neuroanatomie fliegender Reptilien und Auswirkungen auf Flug, Haltung und Verhalten”. Natur. 425 (6961): 950–53. Bibcode:2003Natur.425..950W. doi:10.1038 / nature02048. PMID 14586467. S2CID 4431861.
- ^ “Pterosaur.net :: Ursprünge und Beziehungen”. pterosaur.net. Abgerufen 2020-02-01.
- ^ Andres, Brian; Clark, James M.; Xing, Xu (29.01.2010). “Ein neuer Rhamphorhynchid-Flugsaurier aus dem Oberjura von Xinjiang, China, und die phylogenetischen Beziehungen basaler Flugsaurier” (PDF). Journal of Vertebrate Paleontology. 30 (1): 163–87. doi:10.1080 / 02724630903409220. ISSN 0272-4634. S2CID 53688256.
- ^ Witton, Mark P.; Martill, David M.; Loveridge, Robert F. (2010). “Clipping the Wings of Giant Pterosaurier: Kommentare zu Wingspan Estimations and Diversity”. Acta Geoscientica Sinica. 31: 79–81.
- ^ ein b c d e f G Unwin, David M. (2006). Die Flugsaurier: Aus der tiefen Zeit. New York: Pi Press. p. 246. ISBN 978-0-13-146308-0.
- ^ ein b Frey E, Martill DM (1998). “Weichteilkonservierung in einer Probe von Pterodactylus kochi (Wagner) aus dem Oberjura Deutschlands “. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen. 210 (3): 421–41. doi:10.1127 / njgpa / 210/1998/421.
- ^ ein b c d Naish D, Martill DM (2003). “Flugsaurier – eine erfolgreiche Invasion des prähistorischen Himmels”. Biologe. 50 (5): 213–16.
- ^ ein b Czerkas, SA und Ji, Q. (2002). Ein neuer Rhamphorhynchoid mit einem Kopfkamm und komplexen integumentären Strukturen. In: Czerkas, SJ (Hrsg.). Gefiederte Dinosaurier und der Ursprung der Flucht. Das Dinosaurier-Museum: Blanding, Utah, 15–41. ISBN 1-932075-01-1.
- ^ S. Christopher Bennett (1994). “Taxonomie und Systematik des Flugsauriers der späten Kreidezeit Pteranodon (Pterosauria, Pterodactyloidea). Gelegenheitspapiere des Natural History Museum der University of Kansas. 169: 1–70.
- ^ Bennett SC (2000). “Flugsaurierflug: Die Rolle von Actinofibrillen in der Flügelfunktion”. Historische Biologie. 14 (4): 255–84. doi:10.1080 / 10292380009380572. S2CID 85185457.
- ^ ein b c Kellner, AWA; Wang, X.; Tischlinger, H.; Campos, D.; Hone, DWE; Meng, X. (2009). “Das Weichgewebe von Jeholopterus (Pterosauria, Anurognathidae, Batrachognathinae) und die Struktur der Pterosaurier-Flügelmembran “. Verfahren der Royal Society B.. 277 (1679): 321–29. doi:10.1098 / rspb.2009.0846. PMC 2842671. PMID 19656798.
- ^ ein b Claessens LP, O’Connor PM, Unwin DM (2009). Sereno P (Hrsg.). “Die Entwicklung der Atemwege erleichterte den Ursprung des Flugsaurierfluges und des Luftgigantismus.”. PLUS EINS. 4 (2): e4497. Bibcode:2009PLoSO … 4.4497C. doi:10.1371 / journal.pone.0004497. PMC 2637988. PMID 19223979.
- ^ ein b Unwin DM, Bakhurina NN (1994). “”Sordes pilosus und die Art des Flugsaurier-Flugapparats “. Natur. 371 (6492): 62–64. doi:10.1038 / 371062a0. S2CID 4314989.
- ^ Wang X, Zhou Z, Zhang F, Xu X (2002). “Ein fast vollständig artikulierter Rhamphorhynchoid-Flugsaurier mit außergewöhnlich gut erhaltenen Flügelmembranen und” Haaren “aus der Inneren Mongolei im Nordosten Chinas”. Chinesisches Wissenschaftsbulletin. 47 (3): 3. Bibcode:2002ChSBu..47..226W. doi:10.1360 / 02tb9054. S2CID 86641794.
- ^ Frey, E.; Tischlinger, H.; Buchy, M.-C.; Martill, DM (2003). “Neue Exemplare der Pterosauria (Reptilia) mit Weichteilen mit Auswirkungen auf die Anatomie und Fortbewegung der Pterosaurier”. Geological Society, London, Sonderpublikationen. 217: 233–66. doi:10.1144 / GSL.SP.2003.217.01.14. S2CID 130462931.
- ^ Dyke GJ, Nudds RL, Rayner JM (Juli 2006). “Gliedmaßenunterschiede und Flügelform bei Flugsauriern”. J. Evol. Biol. 19 (4): 1339–42. doi:10.1111 / j.1420-9101.2006.01096.x. PMID 16780534. S2CID 30516133.
- ^ Wilkinson MT, Unwin DM, Ellington CP (2006). “High-Lift-Funktion des Pteroidknochens und Vornähen von Pterosauriern”. Verfahren der Royal Society B.. 273 (1582): 119–26. doi:10.1098 / rspb.2005.3278. PMC 1560000. PMID 16519243.
- ^ Bennett SC (2007). “Artikulation und Funktion des Pteroidknochens von Pterosauriern”. Journal of Vertebrate Paleontology. 27 (4): 881–91. doi:10.1671 / 0272-4634 (2007) 27[881:AAFOTP]2.0.CO; 2.
- ^ Peters D (2009). “Eine Neuinterpretation der Pteroidartikulation bei Pterosauriern”. Journal of Vertebrate Paleontology. 29 (4): 1327–30. doi:10.1671 / 039.029.0407. S2CID 85243363.
- ^ Zhou, Chang-Fu; Schoch, Rainer R. (2011). “Neues Material des nicht-pterodaktyloiden Flugsauriers Changchengopterus pani Lü, 2009 aus der späten Jura-Tiaojishan-Formation des westlichen Liaoning”. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen. 260 (3): 265–75. doi:10.1127 / 0077-7749 / 2011/0131.
- ^ Wang, Xiao-Lin; Kellner, Alexander WA; Jiang, Shun-Xing; Cheng, Xin; Meng, Xi; Rodrigues, Taissa (2010). “Neue Langschwanz-Flugsaurier (Wukongopteridae) aus West-Liaoning, China”. Anais da Academia Brasileira de Ciências. 82 (4): 1045–62. doi:10.1590 / s0001-37652010000400024. PMID 21152776.
- ^ Wilkinson MT; Unwin DM; Ellington CP (2006). “High-Lift-Funktion des Pteroidknochens und Vornieren von Pterosauriern”. Verfahren der Royal Society B.. 273 (1582): 119–26. doi:10.1098 / rspb.2005.3278. PMC 1560000. PMID 16519243.
- ^ ein b c d Witton MP, Naish D (2008). McClain CR (Hrsg.). “Eine Neubewertung der funktionellen Morphologie und Paläoökologie des Pterosauriers der Azhdarchiden”. PLUS EINS. 3 (5): e2271. Bibcode:2008PLoSO … 3.2271W. doi:10.1371 / journal.pone.0002271. PMC 2386974. PMID 18509539.
- ^ Goldfuss, A (1831). “Beiträge zur Erkentniss diversner Reptilien der Vorwelt”. Nova Acta Academiae Leopoldinae. 15: 61–128.
- ^ Benton, Michael J.; Xu, Xing; Orr, Patrick J.; Kaye, Thomas G.; Pittman, Michael; Kearns, Stuart L.; McNamara, Maria E.; Jiang, Baoyu; Yang, Zixiao (Januar 2019). “Pterosaurier-Integumentarstrukturen mit komplexen federartigen Verzweigungen”. Naturökologie & Evolution. 3 (1): 24–30. doi:10.1038 / s41559-018-0728-7. hdl:1983 / 1f7893a1-924d-4cb3-a4bf-c4b1592356e9. ISSN 2397-334X. PMID 30568282. S2CID 56480710.
- ^ Briggs, Helen (2018-12-17). “Fell fliegt über neue Flugsaurierfossilien”. BBC News. Abgerufen 2018-12-19.
- ^ Unwin & Martill (28. September 2020). “Keine Protofedern an Flugsauriern”. Naturökologie und Evolution. doi:10.1038 / s41559-020-01308-9. PMID 32989266.
- ^ Kellner; et al. (2009). “Das Weichgewebe von Jeholopterus (Pterosauria, Anurognathidae, Batrachognathinae) und die Struktur der Pterosaurierflügelmembran”. ResearchGate: 321–29.
- ^ Unwin, David M.; Bakhurina, Natasha N. (September 1994). “Sordes pilosus und die Natur des Flugsaurier-Flugapparats”. Natur. 371 (6492): 62–64. doi:10.1038 / 371062a0. ISSN 1476-4687. S2CID 4314989.
- ^ Yang, Zixiao; Jiang, Baoyu; McNamara, Maria E.; Kearns, Stuart L.; Pittman, Michael; Kaye, Thomas G.; Orr, Patrick J.; Xu, Xing; Benton, Michael J. (28.09.2020). “Antwort an: Keine Protofedern an Flugsauriern”. Naturökologie & Evolution: 1–2. doi:10.1038 / s41559-020-01309-8. ISSN 2397-334X. PMID 32989267.
- ^ Collini, CA (1784). “Sur quelques Zoolithes du Cabinet d’Histoire naturelle de SASE Palatine & de Bavière, à Mannheim.” Acta Theodoro-Palatinae Mannheim 5 Pars PhysicaS. 58–103 (1 Platte).
- ^ Wagler, J. (1830). Natürliches System der Amphibien München, 1830: 1–354.
- ^ Cuvier G (1801). “”[Reptile volant]. In: Extrait d’un ouvrage sur les espèces de quadrupèdes nicht auf einem trouvé les ossemens dans l’intérieur de la terre “. Journal de Physique, de Chimie und d’Histoire Naturelle. 52: 253–67.
- ^ Cuvier, G., 1809, “Mémoire sur le squelette fossile d’un Reptil volant des environs d’Aichstedt, que quelques naturalistes ont pris pour un oiseau und donc nous formons un genre de Sauriens, sous le nom de Ptero-Dactyle” , Annales du Musée d’Histoire Naturelle, Paris, 13 S. 424–37
- ^ Rafinesque, CS, 1815, Analysieren Sie die Natur oder das Tableau der Universitäten und des Korps, Palermo
- ^ Von Soemmerring, ST, 1812 der Naturalien-Sammlung der politischen Akademie der Wissenschaften zu Münchensicht “, Denkschriften der politischen bayerischen Akademie der Wissenschaften, München: mathematisch-physikalische Klasse 3: 89–158
- ^ Newman, E (1843). “Anmerkung zum Pterodactyle-Stamm, der als Beuteltierfledermäuse gilt”. Zoologe. 1: 129–31.
- ^ Kaup, J. (1834). “Versuch einer Einheilung der Todesethiere in 6 Bezirken und der Amphibien in 6 Ordnungen”. Isis von Oken. 1834: 315.
- ^ Seeley, HG, 1870, Ornithosauria – eine elementare Untersuchung der Knochen von Pterodaktylen, Cambridge University Press
- ^ Seeley, HG, 1901, Drachen der Luft: Ein Bericht über ausgestorbene fliegende Reptilien, London: Methuen
- ^ Mivart, G. (1881). “Ein beliebter Bericht über Chamaäleons”. Natur. 24: 309–38.
- ^ Marsh, OC (1882). “Die Flügel der Pterodaktylen”. American Journal of Science. 3 (16): 223.
- ^ Zittel, KA (1882). “Über Flugsaurier aus dem lithgesteuerten Schiefer Bayerns”. Palaeonthographica. 29: 47–80.
- ^ Broili, F., 1927, “Ein Ramphorhynchus mit Spuren von Haarbedeckung”, Sitzungsberichte der Bayerischen Akademie der Wissenschaften p. 49-67
- ^ Edinger, T. (1927). “Das Gehirn der Flugsaurier” (PDF). Zeitschrift für Anatomie und Entwicklungsgeschichte. 83 (1/3): 105–12. doi:10.1007 / bf02117933. S2CID 19084773.
- ^ Hankin EH & Watson DSM; “Auf dem Flug der Pterodaktylen”, Das LuftfahrtjournalOktober 1914, S. 324–35
- ^ Bakker, Robert, 1986, Die Dinosaurier-Häresien, Londen: Penguin Books, 1988, p. 283
- ^ Padian, K. (1979). “Die Flügel der Flugsaurier: Ein neuer Look”. Entdeckung. 14: 20–29.
- ^ Padian, K., 1980, Untersuchungen zur Struktur, Entwicklung und Flucht von Flugsauriern (Reptilien: Flugsaurier), Ph.D. Diss., Institut für Biologie, Yale University
- ^ ein b Padian K (1983). “Eine Funktionsanalyse des Fliegens und Gehens bei Flugsauriern”. Paläobiologie. 9 (3): 218–39. doi:10.1017 / S009483730000765X. JSTOR 2400656.
- ^ Wellnhofer, P., 1978, Handbuch der Paläoherpetologie XIX. Flugsaurier, Urban & Fischer, München
- ^ Dyke, GJ McGowan; Nudds, RL; Smith, D. (2009). “Die Form der Pterosaurier-Evolution: Beweise aus dem Fossilienbestand”. Zeitschrift für Evolutionsbiologie. 22 (4): 890–98. doi:10.1111 / j.1420-9101.2008.01682.x. PMID 19210587. S2CID 32518380.
- ^ ein b Witton, Mark (2013). Flugsaurier: Naturgeschichte, Evolution, Anatomie. Princeton University Press. ISBN 978-0691150611.
- ^ Benton, MJ (1999). “”Scleromochlus taylori und der Ursprung der Dinosaurier und Flugsaurier “. Philosophische Transaktionen der Royal Society B: Biologische Wissenschaften. 354 (1388): 1423–46. doi:10.1098 / rstb.1999.0489. PMC 1692658.
- ^ Bennett, S. Christopher (1996). “Die phylogenetische Position der Pterosauria innerhalb der Archosauromorpha”. Zoologisches Journal der Linnean Society. 118 (3): 261–308. doi:10.1111 / j.1096-3642.1996.tb01267.x.
- ^ Irmis, RB; Nesbitt, SJ; Padian, K.; Smith, ND; Turner, AH; Woody, D.; Downs, A. (2007). “Eine spättriassische Dinosauromorph-Assemblage aus New Mexico und der Aufstieg der Dinosaurier”. Wissenschaft. 317 (5836): 358–61. Bibcode:2007Sci … 317..358I. doi:10.1126 / science.1143325. PMID 17641198. S2CID 6050601.
- ^ ein b Hone DWE; Benton MJ (2007). “Eine Bewertung der phylogenetischen Beziehungen der Flugsaurier zu den archosauromorphen Reptilien”. Zeitschrift für systematische Paläontologie. 5 (4): 465–69. doi:10.1017 / S1477201907002064. S2CID 86145645.
- ^ Nesbitt, SJ (2011). “Die frühe Entwicklung der Archosaurier: Beziehungen und der Ursprung der Hauptkladen” (PDF). Bulletin des American Museum of Natural History. 352: 1–292. doi:10.1206 / 352.1. hdl:2246/6112. S2CID 83493714.
- ^ Ezcurra, Martín D. (28.04.2016). “Die phylogenetischen Beziehungen basaler Archosauromorphe mit Schwerpunkt auf der Systematik proterosuchischer Archosauriformen”. PeerJ. 4: e1778. doi:10.7717 / peerj.1778. ISSN 2167-8359. PMC 4860341. PMID 27162705.
- ^ Bennett, SC (2020). “Neubewertung der Trias-Archosauriform Scleromochlus taylori: weder Läufer noch Zweibeiner, sondern Trichter “. PeerJ. 8: e8418. doi:10.7717 / peerj.8418. PMID 32117608.
- ^ Rupert Wild, 1983, “Über die Ursprung der Flugsaurier”, Weltenberger Akademie, Erwin Rutte-FestschriftS. 231–38
- ^ ein b c d Witton, Mark P. (2015). “Waren frühe Flugsaurier unfähige Landlokomotoren?”. PeerJ. 3: e1018. doi:10.7717 / peerj.1018. PMC 4476129. PMID 26157605.
- ^ BBC-Dokumentarfilm: Mit Dinosauriern spazieren gehen (Folge 4) – Riese des Himmels bei 22 ‘, Tim Haines, 1999
- ^ Slack KE, Jones CM, Ando T. et al. (Juni 2006). “Frühe Pinguinfossilien sowie mitochondriale Genome kalibrieren die Vogelentwicklung”. Molekularbiologie und Evolution. 23 (6): 1144–55. doi:10.1093 / molbev / msj124. PMID 16533822.
- ^ Butler, Richard J.; Barrett, Paul M.; Nowbath, Stephen & Upchurch, Paul (2009). “Abschätzung der Auswirkungen von Stichprobenverzerrungen auf die Diversitätsmuster von Flugsauriern: Auswirkungen auf Hypothesen des kompetitiven Ersatzes von Vögeln und Flugsauriern”. Paläobiologie. 35 (3): 432–46. doi:10.1666 / 0094-8373-35.3.432. S2CID 84324007.
- ^ ein b Longrich, NR; Martill, DM; Andres, B. (2018). “Spätmaastrichtische Flugsaurier aus Nordafrika und Massensterben von Flugsauriern an der Grenze zwischen Kreide und Paläogen”. PLOS Biologie. 16 (3): e2001663. doi:10.1371 / journal.pbio.2001663. PMC 5849296. PMID 29534059.
- ^ Andres, B.; Myers, TS (2013). “Lone Star Pterosaurier”. Erd- und Umweltwissenschaftliche Transaktionen der Royal Society of Edinburgh. 103 (3–4): 1. doi:10.1017 / S1755691013000303.
- ^ Agnolin, Federico L. & Varricchio, David (2012). “Systematische Neuinterpretation von Piksi barbarulna Varricchio, 2002 aus der Two Medicine Formation (Oberkreide) im Westen der USA (Montana) als Flugsaurier statt als Vogel “ (PDF). Geodiversitas. 34 (4): 883–94. doi:10.5252 / g2012n4a10. S2CID 56002643.
- ^ Prondvai, E.; Bodor, ER; Ösi, A. (2014). “Spiegelt die Morphologie die auf Osteohistologie basierende Ontogenese wider? Eine Fallstudie von Kiefer-Symphysen aus der späten Kreidezeit des Flugsauriers aus Ungarn enthüllt verborgene taxonomische Vielfalt.” (PDF). Paläobiologie. 40 (2): 288–321. doi:10.1666 / 13030. S2CID 85673254.
- ^ Martin-Silverstone, Elizabeth; Witton, Mark P.; Arbor, Victoria M.; Currie, Philip J. (2016). “Ein kleiner azhdarchoider Flugsaurier aus der jüngsten Kreidezeit, dem Zeitalter der fliegenden Riesen”. Royal Society Open Science. 3 (8): 160333. Bibcode:2016RSOS …. 360333M. doi:10.1098 / rsos.160333. PMC 5108964. PMID 27853614.
- ^ Haluza, A.; Apesteguía, S. (2007). “Flugsaurierreste (Archosauria, Ornithodira) aus der frühen Spätkreide von” La Buitrera “, Río Negro, Argentinien”. XXIII Jornadas Argentinas de Paleontología de Vertebrados.
- ^ Kellner, AW (2003). “Pterosaurier-Phylogenie und Kommentare zur Evolutionsgeschichte der Gruppe”. Geological Society, London, Sonderpublikationen. 217 (1): 105–37. Bibcode:2003GSLSP.217..105K. doi:10.1144 / gsl.sp.2003.217.01.10. S2CID 128892642.
- ^ Nesbitt, SJ, Desojo, JB & Irmis, RB (2013). Anatomie, Phylogenie und Paläobiologie der frühen Archosaurier und ihrer Verwandten. Geologische Gesellschaft von London. ISBN 1862393613
- ^ de Queiroz. K., Cantino. PD, Gauthier. JA eds. (2020). Phylonyme: Ein Begleiter des PhyloCodes. CRC Press Boca Raton, FL
- ^ Padian, K. (1997). “Pterosauromorpha”, S. 617–18 in Currie, PJ und Padian, K. Die Enzyklopädie der Dinosaurier. Akademische Presse. ISBN 0122268105.
- ^ Lü J.; Unwin DM; Xu L.; Zhang X. (2008). “Ein neuer azhdarchoider Flugsaurier aus der Unterkreide Chinas und seine Auswirkungen auf die Phylogenie und Evolution der Flugsaurier”. Naturwissenschaften. 95 (9): 891–97. Bibcode:2008NW ….. 95..891L. doi:10.1007 / s00114-008-0397-5. PMID 18509616. S2CID 13458087.
- ^ ein b Alleyne, Richard (1. Oktober 2008). “Pterodaktylen waren zu schwer zum Fliegen, behaupten Wissenschaftler”. Der Telegraph. Abgerufen 2. März 2012.
- ^ Powell, Devin (2. Oktober 2008). “Waren Flugsaurier zu groß zum Fliegen?”. NewScientist. Abgerufen 2. März 2012.
- ^ Templin, RJ; Chatterjee, Sankar (2004). Haltung, Fortbewegung und Paläoökologie von Flugsauriern. Boulder, Colorado: Geologische Gesellschaft von Amerika. p. 60. ISBN 978-0-8137-2376-1.
- ^ Naish, Darren (18. Februar 2009). “Flugsaurier atmeten vogelartig und hatten aufblasbare Luftsäcke in den Flügeln.”. ScienceBlogs. Archiviert vom Original am 21. Februar 2009. Abgerufen 3. April 2016.
- ^ ein b c “Warum Flugsaurier doch nicht so gruselig waren”. Die Zeitung Observer. 11. August 2013. Abgerufen 12. August 2013.
- ^ ein b Hecht, Jeff (16. November 2010). “Haben sich riesige Flugsaurier wie Vampirfledermäuse hochgewölbt?”. NewScientist. Abgerufen 2. März 2012.
- ^ MacCready, P. (1985). “Das große Pterodaktylus-Projekt” (PDF). Ingenieurwesen & Wissenschaft, California Institute of Technology. 49 (2): 18–24.
- ^ Molotsky, Irvin (28. Januar 1986). “Mit flatternden Flügeln fliegt das Modell pterodaktisch in die Luft”. New York Times.
- ^ Geist, N.; Hillenius, W.; Frey, E.; Jones, T.; Elgin, R. (2014). “Atmen in einer Box: Einschränkungen der Lungenbeatmung bei riesigen Flugsauriern”. Die anatomische Aufzeichnung. 297 (12): 2233–53. doi:10.1002 / ar.22839. PMID 24357452. S2CID 27659270.
- ^ Hopson JA (1977). “Relative Gehirngröße und Verhalten bei archosaurischen Reptilien”. Jahresrückblick auf Ökologie und Systematik. 8: 429–48. doi:10.1146 / annurev.es.08.110177.002241.
- ^ Anthes, Emily (18. November 2013). “Kaltblütig bedeutet nicht dumm” – über NYTimes.com.
- ^ Codorniú, Laura; Paulina Carabajal, Ariana; Pol, Diego; Unwin, David; Rauhut, Oliver WM (2016). “Ein Jura-Flugsaurier aus Patagonien und der Ursprung des Pterodaktyloid-Neurokraniums”. PeerJ. 4: e2311. doi:10.7717 / peerj.2311. PMC 5012331. PMID 27635315.
- ^ Padian K (2003). “Pterosaurier Haltung und Gang und die Interpretation von Spuren”. Ichnos. 10 (2–4): 115–26. doi:10.1080 / 10420940390255501. S2CID 129113446.
- ^ Hwang K., Huh M., Lockley MG, Unwin DM, Wright JL (2002). “Neue Flugsaurierspuren (Pteraichnidae) aus der spätkreidezeitlichen Uhangri-Formation im Südwesten Koreas”. Geologisches Magazin. 139 (4): 421–35. Bibcode:2002GeoM..139..421H. doi:10.1017 / S0016756802006647. S2CID 54996027.
- ^ Unwin DM (1997). “Flugsaurier-Spuren und die terrestrische Fähigkeit von Flugsauriern”. Lethaia. 29 (4): 373–86. doi:10.1111 / j.1502-3931.1996.tb01673.x.
- ^ ein b c Witton, Mark (2013). Flugsaurier: Naturgeschichte, Evolution, Anatomie. Princeton University Press. p. 51. ISBN 978-0691150611.
- ^ Longrich, NR; Martill, DM; Andres, B. (2018). “Spätmaastrichtische Flugsaurier aus Nordafrika und Massensterben von Flugsauriern an der Grenze zwischen Kreide und Paläogen”. PLOS Biologie. 16 (3): e2001663. doi:10.1371 / journal.pbio.2001663. PMC 5849296. PMID 29534059.
- ^ Bennett, SC (2007). “Ein zweites Exemplar des Flugsauriers Anurognathus ammoni“. Paläontologische Zeitschrift. 81 (4): 376–98. doi:10.1007 / bf02990250. S2CID 130685990.
- ^ Andres, B.; Clark, JM; Xing, X. (2010). “Ein neuer Rhamphorhynchid-Flugsaurier aus dem Oberjura von Xinjiang, China, und die phylogenetischen Beziehungen basaler Flugsaurier” (PDF). Journal of Vertebrate Paleontology. 30 (1): 163–87. doi:10.1080 / 02724630903409220. S2CID 53688256.
- ^ Lü J.; Xu L.; Chang H.; Zhang X. (2011). “Ein neuer darwinopterider Flugsaurier aus dem Mittleren Jura des westlichen Liaoning im Nordosten Chinas und seine ökologischen Auswirkungen”. Acta Geologica Sinica – Englische Ausgabe. 85 (3): 507–14. doi:10.1111 / j.1755-6724.2011.00444.x.
- ^ Wu, Wen-Hao; Zhou, Chang-Fu; Andres, Brian (2017). “Der zahnlose Flugsaurier Jidapterus edentus (Pterodactyloidea: Azhdarchoidea) aus der frühen Kreidezeit Jehol Biota und seine paläoökologischen Auswirkungen “. PLUS EINS. 12 (9): e0185486. Bibcode:2017PLoSO..1285486W. doi:10.1371 / journal.pone.0185486. PMC 5614613. PMID 28950013.
- ^ Pêgas, RV & Kellner, AW (2015). Vorläufige myologische Rekonstruktion des Unterkiefers von Thalassodromeus sethi (Pterodactyloidea: Tapejaridae). Flugsaurier 2015 Portsmouth, Abstracts, 47–48
- ^ Witton, MP; Naish, D. (2015). “Azhdarchid Pterosaurier: Pelikanimitationen mit Wasserschleppnetzen oder” terrestrische Stalker “?”. Acta Palaeontologica Polonica. 60 (3). doi:10.4202 / app.00005.2013.
- ^ Naish, D.; Witton, MP (2017). “Die Halsbiomechanik zeigt, dass riesige siebenbürgische Azhdarchiden-Flugsaurier kurzhalsige Bogenräuber waren.”. PeerJ. 5: e2908. doi:10.7717 / peerj.2908. PMC 5248582. PMID 28133577.
- ^ Witton, M.; Brusatte, S.; Dyke, G.; Naish, D.; Norell, M.; Vremir, M. (2013). Flugsaurier-Oberherren von Siebenbürgen: kurzhalsige Riesenazhdarchiden in der späten Kreidezeit Rumäniens. Das jährliche Symposium für Paläontologie und vergleichende Anatomie von Wirbeltieren. Edinburgh. Archiviert von das Original am 06.04.2016.
- ^ Martill, DM; Ibrahim, N. (2015). “Eine ungewöhnliche Modifikation der Kiefer in vgl. Alanqa, ein azhdarchidischer Flugsaurier aus der Kreidezeit aus den Kem Kem-Betten Marokkos “ (PDF). Kreideforschung. 53: 59–67. doi:10.1016 / j.cretres.2014.11.001.
- ^ Martill, David M.; Smith, Roy E.; Longrich, Nicholas; Brown, James (2020-09-03). “Hinweise auf taktile Fütterung bei Flugsauriern: eine empfindliche Spitze des Schnabels von Lonchodraco giganteus (Pterosauria, Lonchodectidae) aus der Oberkreide im Süden Englands”. Kreideforschung: 104637. doi:10.1016 / j.cretres.2020.104637. ISSN 0195-6671.
- ^ Buffetaut E, Martill D, Escuillié F (Juli 2004). “Flugsaurier als Teil einer Spinosaurierdiät”. Natur. 430 (6995): 33. Bibcode:2004Natur.429 … 33B. doi:10.1038 / 430033a. PMID 15229562. S2CID 4398855.
- ^ Ji Q, Ji SA, Cheng YN et al. (Dezember 2004). “Paläontologie: Flugsaurier-Ei mit einer ledrigen Schale”. Natur. 432 (7017): 572. doi:10.1038 / 432572a. PMID 15577900. S2CID 4416203.
- ^ Lü J.; Unwin DM; Deeming DC; Jin X.; Liu Y.; Ji Q. (2011). “Eine Ei-Erwachsenen-Assoziation, Geschlecht und Fortpflanzung bei Flugsauriern”. Wissenschaft. 331 (6015): 321–24. doi:10.1126 / science.1197323. PMID 21252343. S2CID 206529739.
- ^ ein b Wang, Xiaolin (2014). “Sexuell dimorphe dreidimensional konservierte Flugsaurier und ihre Eier aus China”. Aktuelle Biologie. 24 (12): 1323–30. doi:10.1016 / j.cub.2014.04.054. PMID 24909325.[permanent dead link]
- ^ Grellet-Tinner G, Wroe S., Thompson MB, Ji Q (2007). “Ein Hinweis zum Verschachtelungsverhalten von Flugsauriern”. Historische Biologie. 19 (4): 273–77. doi:10.1080 / 08912960701189800. S2CID 85055204.
- ^ Xiaolin Wang, Kellner Alexander WA; Cheng, Xin; Jiang, Shunxing; Wang, Qiang; Sayão Juliana, M.; Rordrigues Taissa, Costa Fabiana R.; Beschichtung; Meng, Xi; Zhou, Zhonghe (2015). “Eierschale und Histologie geben Einblick in die Lebensgeschichte eines Flugsauriers mit zwei funktionellen Eierstöcken”. Anais da Academia Brasileira de Ciências. 87 (3): 1599–1609. doi:10.1590 / 0001-3765201520150364. PMID 26153915.
- ^ Wang X, Zhou Z (Juni 2004). “Paläontologie: Flugsaurierembryo aus der frühen Kreidezeit”. Natur. 429 (6992): 621. Bibcode:2004Natur.429..621W. doi:10.1038 / 429621a. PMID 15190343. S2CID 4428545.
- ^ ein b “Pterosaurier-Jungtiere brauchten ihre Eltern, Eierschatz enthüllt (Update)”. phys.org. Abgerufen 2020-03-21.
- ^ ein b c Bennett SC (1995). “Eine statistische Studie von Rhamphorhynchus aus dem Solnhofener Kalkstein Deutschlands: Jahresklassen einer einzigen großen Art “. Zeitschrift für Paläontologie. 69 (3): 569–80. doi:10.1017 / S0022336000034946. JSTOR 1306329.
- ^ ein b Prondvai, E.; Stein, K.; Ősi, A.; Sander, MP (2012). Soares, Daphne (Hrsg.). “Lebensgeschichte von Rhamphorhynchus abgeleitet aus der Knochenhistologie und der Vielfalt der Wachstumsstrategien der Flugsaurier “. PLUS EINS. 7 (2): e31392. doi:10.1371 / journal.pone.0031392. PMC 3280310. PMID 22355361.
- ^ “Erste 3D-Flugsaurier-Eier, die bei ihren Eltern gefunden wurden”. phys.org. Abgerufen 2020-03-21.
- ^ Unwin, David Michael; Deeming, D. Charles (2019). “Pränatale Entwicklung bei Flugsauriern und ihre Auswirkungen auf ihre postnatale Bewegungsfähigkeit”. Verfahren der Royal Society B: Biological Sciences. 286 (1904). doi:10.1098 / rspb.2019.0409. PMC 6571455. PMID 31185866.
- ^ Schmitz, L.; Motani, R. (2011). “Nächtlichkeit bei Dinosauriern, abgeleitet aus der Morphologie des Sklerarings und der Umlaufbahn”. Wissenschaft. 332 (6030): 705–08. Bibcode:2011Sci … 332..705S. doi:10.1126 / science.1200043. PMID 21493820. S2CID 33253407.
- ^ ein b c Hone, D. (2010). “Flugsaurier in der Populärkultur.” Pterosaur.net, Zugriff am 27. August 2010.
- ^ Frey, E., Martill, D. und Buchy, M. (2003). “Ein neues Ornithocheirid mit Haube aus der Unterkreide im Nordosten Brasiliens und der ungewöhnliche Tod eines ungewöhnlichen Flugsauriers” in: Buffetaut, E., und Mazin, J.-M. (Hrsg.). Evolution und Paläobiologie der Flugsaurier. Sonderpublikation der Geologischen Gesellschaft 217: 56–63. ISBN 1-86239-143-2.
- ^ Gonzales, Dave (12. Oktober 2016). “Eine Zusammenstellung aller verrückten ‘Godzilla’-Filme in Monstergröße”. Thrillist. Abgerufen 11. Juli 2019.;; Sharf, Zack (10. Dezember 2018). “”‘Godzilla: Der Trailer des Königs der Monster verwandelt Mothra, Rodan und mehr in ein episches Spektakel “ (Video). IndieWire. Abgerufen 11. Juli 2019.
- ^ “Mythen & Missverständnisse”. Pterosaur.net
Zitierte Werke[edit]
- Berry, Mark F. (2005). Die Dinosaurier-Filmographie. McFarland & Company. ISBN 978-0786424535.
- Wellnhofer, Peter (1991). Die illustrierte Enzyklopädie der Flugsaurier: Eine illustrierte Naturgeschichte der fliegenden Reptilien des Mesozoikums. Halbmondbücher. ISBN 0517037017.
- Witton, Mark (2013). Flugsaurier: Naturgeschichte, Evolution, Anatomie. Princeton University Press. ISBN 978-0691150611.
Externe Links[edit]
| Wikisource enthält mehrere Originaltexte zu folgenden Themen: Flugsaurier |
Recent Comments