Ökosystemökologie – Wikipedia
Die Untersuchung lebender und nicht lebender Komponenten von Ökosystemen und ihrer Wechselwirkungen


Ökosystemökologie ist die integrierte Untersuchung lebender (biotischer) und nicht lebender (abiotischer) Komponenten von Ökosystemen und ihrer Wechselwirkungen innerhalb eines Ökosystemrahmens. Diese Wissenschaft untersucht, wie Ökosysteme funktionieren, und bezieht dies auf ihre Bestandteile wie Chemikalien, Grundgestein, Boden, Pflanzen und Tiere.
Die Ökosystemökologie untersucht physikalische und biologische Strukturen und untersucht, wie diese Ökosystemmerkmale miteinander interagieren. Dies hilft uns letztendlich zu verstehen, wie qualitativ hochwertiges Wasser und eine wirtschaftlich tragfähige Rohstoffproduktion aufrechterhalten werden können. Ein Hauptaugenmerk der Ökosystemökologie liegt auf funktionalen Prozessen, ökologischen Mechanismen, die die Struktur und die Dienstleistungen von Ökosystemen erhalten. Dazu gehören Primärproduktivität (Produktion von Biomasse), Zersetzung und trophische Wechselwirkungen.
Studien zur Funktion des Ökosystems haben das Verständnis des Menschen für eine nachhaltige Produktion von Futter, Ballaststoffen, Brennstoffen und die Bereitstellung von Wasser erheblich verbessert. Funktionale Prozesse werden durch Klima, Störung und Management auf regionaler bis lokaler Ebene vermittelt. Die Ökosystemökologie bietet somit einen leistungsstarken Rahmen für die Identifizierung ökologischer Mechanismen, die mit globalen Umweltproblemen, insbesondere der globalen Erwärmung und dem Abbau von Oberflächenwasser, interagieren.
Dieses Beispiel zeigt einige wichtige Aspekte von Ökosystemen:
- Ökosystemgrenzen sind oft nebulös und können zeitlich schwanken
- Organismen in Ökosystemen sind abhängig von biologischen und physikalischen Prozessen auf Ökosystemebene
- Benachbarte Ökosysteme interagieren eng miteinander und sind häufig voneinander abhängig, um die Gemeinschaftsstruktur und funktionale Prozesse aufrechtzuerhalten, die die Produktivität und die biologische Vielfalt erhalten
Diese Eigenschaften führen auch zu praktischen Problemen beim Management natürlicher Ressourcen. Wer wird welches Ökosystem verwalten? Wird das Fällen von Holz im Wald die Freizeitfischerei im Bach beeinträchtigen? Diese Fragen sind für Landverwalter schwer zu beantworten, während die Grenze zwischen den Ökosystemen unklar bleibt. obwohl Entscheidungen in einem Ökosystem das andere beeinflussen. Wir brauchen ein besseres Verständnis der Wechselwirkungen und Abhängigkeiten dieser Ökosysteme und der Prozesse, die sie aufrechterhalten, bevor wir beginnen können, diese Fragen zu beantworten.
Die Ökosystemökologie ist ein inhärent interdisziplinäres Forschungsgebiet. Ein individuelles Ökosystem besteht aus Populationen von Organismen, die innerhalb von Gemeinschaften interagieren und zum Nährstoffkreislauf und zum Energiefluss beitragen. Das Ökosystem ist die Hauptstudieneinheit in der Ökosystemökologie.
Bevölkerung, Gemeinschaft und physiologische Ökologie liefern viele der zugrunde liegenden biologischen Mechanismen, die die Ökosysteme und die von ihnen aufrechterhaltenen Prozesse beeinflussen. Der Energiefluss und der Stoffkreislauf auf Ökosystemebene werden in der Ökosystemökologie häufig untersucht. Insgesamt wird diese Wissenschaft jedoch eher nach Themen als nach Maßstäben definiert. Die Ökosystemökologie betrachtet Organismen und abiotische Energie- und Nährstoffpools als ein integriertes System, das sie von assoziierten Wissenschaften wie der Biogeochemie unterscheidet.[1]
Biogeochemie und Hydrologie konzentrieren sich auf verschiedene grundlegende Ökosystemprozesse wie den biologisch vermittelten chemischen Kreislauf von Nährstoffen und den physikalisch-biologischen Kreislauf von Wasser. Die Ökosystemökologie bildet die mechanistische Grundlage für regionale oder globale Prozesse, die von der Hydrologie von Landschaft zu Region, der globalen Biogeochemie und der Erdsystemwissenschaft umfasst werden.[1]
Geschichte[edit]
Die Ökosystemökologie ist philosophisch und historisch in der terrestrischen Ökologie verwurzelt. Das Ökosystemkonzept hat sich in den letzten 100 Jahren rasant weiterentwickelt, wobei wichtige Ideen von Frederic Clements entwickelt wurden, einem Botaniker, der sich für spezifische Definitionen von Ökosystemen aussprach und dass physiologische Prozesse für deren Entwicklung und Persistenz verantwortlich waren.[2] Obwohl die meisten Definitionen des Clements-Ökosystems zunächst von Henry Gleason und Arthur Tansley und später von zeitgenössischen Ökologen stark überarbeitet wurden, bleibt die Idee, dass physiologische Prozesse für die Struktur und Funktion des Ökosystems von grundlegender Bedeutung sind, für die Ökologie von zentraler Bedeutung.

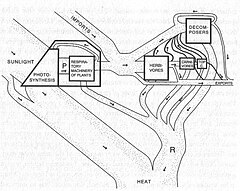
Spätere Arbeiten von Eugene Odum und Howard T. Odum quantifizierten Energie- und Materieflüsse auf Ökosystemebene und dokumentierten so die allgemeinen Ideen von Clements und seinem Zeitgenossen Charles Elton.
In diesem Modell waren die Energieflüsse durch das gesamte System abhängig von biotischen und abiotischen Wechselwirkungen jeder einzelnen Komponente (Spezies, anorganische Nährstoffpools usw.). Spätere Arbeiten zeigten, dass diese Wechselwirkungen und Flüsse auf Nährstoffkreisläufe angewendet wurden, sich im Laufe der Folge veränderten und die Produktivität des Ökosystems stark kontrollierten.[4][5] Energie- und Nährstofftransfers sind ökologischen Systemen angeboren, unabhängig davon, ob sie aquatisch oder terrestrisch sind. So ist die Ökosystemökologie aus wichtigen biologischen Studien von Pflanzen, Tieren, terrestrischen, aquatischen und marinen Ökosystemen hervorgegangen.
Ökosystem-Dienstleistungen[edit]
Ökosystemleistungen sind ökologisch vermittelte Funktionsprozesse, die für die Erhaltung gesunder menschlicher Gesellschaften unerlässlich sind.[6] Wasserversorgung und -filtration, Produktion von Biomasse in der Forstwirtschaft, Landwirtschaft und Fischerei sowie Entfernung von Treibhausgasen wie Kohlendioxid (CO)2) aus der Atmosphäre sind Beispiele für Ökosystemleistungen, die für die öffentliche Gesundheit und wirtschaftliche Chancen von wesentlicher Bedeutung sind. Der Nährstoffkreislauf ist ein Prozess, der für die land- und forstwirtschaftliche Produktion von grundlegender Bedeutung ist.
Wie bei den meisten Ökosystemprozessen ist der Nährstoffkreislauf jedoch kein Ökosystemmerkmal, das auf das wünschenswerteste Niveau „gewählt“ werden kann. Die Maximierung der Produktion in degradierten Systemen ist eine zu vereinfachte Lösung für die komplexen Probleme von Hunger und wirtschaftlicher Sicherheit. Beispielsweise hat der intensive Einsatz von Düngemitteln im Mittleren Westen der USA zu einer Verschlechterung der Fischerei im Golf von Mexiko geführt.[7] Bedauerlicherweise wurde für die Landwirtschaft in Industrie- und Entwicklungsländern eine „Grüne Revolution“ intensiver chemischer Düngung empfohlen.[8][9] Diese Strategien riskieren eine Veränderung von Ökosystemprozessen, die möglicherweise schwer wiederherzustellen sind, insbesondere wenn sie in großem Maßstab ohne angemessene Bewertung der Auswirkungen angewendet werden. Es kann viele Jahre dauern, bis sich Ökosystemprozesse von erheblichen Störungen erholt haben.[5]
Beispielsweise hat die großflächige Waldrodung im Nordosten der Vereinigten Staaten im 18. und 19. Jahrhundert die Bodentextur, die dominierende Vegetation und den Nährstoffkreislauf in einer Weise verändert, die die heutige Waldproduktivität beeinflusst.[10][11] In Verbindung mit Plänen zur Wiederherstellung wesentlicher Prozesse ist eine Einschätzung der Bedeutung der Ökosystemfunktion für die Aufrechterhaltung der Produktivität in der Land- oder Forstwirtschaft erforderlich. Verbesserte Kenntnisse der Ökosystemfunktion werden dazu beitragen, langfristige Nachhaltigkeit und Stabilität in den ärmsten Teilen der Welt zu erreichen.
Betrieb[edit]
Die Produktivität von Biomasse ist eine der offensichtlichsten und wirtschaftlich wichtigsten Ökosystemfunktionen. Die Akkumulation von Biomasse beginnt auf zellulärer Ebene über die Photosynthese. Die Photosynthese erfordert Wasser und folglich korrelieren globale Muster der jährlichen Biomasseproduktion mit dem jährlichen Niederschlag.[12] Die Produktivitätsmengen hängen auch von der Gesamtkapazität der Pflanzen ab, Sonnenlicht einzufangen, das direkt mit der Blattfläche der Pflanzen und dem N-Gehalt korreliert.
Die Netto-Primärproduktivität (KKW) ist das primäre Maß für die Anreicherung von Biomasse in einem Ökosystem. Die Netto-Primärproduktivität kann durch eine einfache Formel berechnet werden, bei der die Gesamtproduktivitätsmenge durch Aufrechterhaltung biologischer Prozesse an die Gesamtproduktivitätsverluste angepasst wird:
- KKW = GPP – R.Produzent

Wobei GPP die Bruttoprimärproduktivität ist und R.Produzent ist Photosynthese (Kohlenstoff), die durch Zellatmung verloren geht.
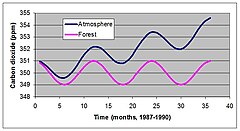
KKW ist schwer zu messen, aber eine neue Technik, die als Wirbel-Co-Varianz bekannt ist, hat Aufschluss darüber gegeben, wie natürliche Ökosysteme die Atmosphäre beeinflussen. Abbildung 4 zeigt saisonale und jährliche Veränderungen des CO2 Konzentration gemessen in Mauna Loa, Hawaii von 1987 bis 1990. CO2 Die Konzentration nahm stetig zu, aber die Abweichungen innerhalb des Jahres waren größer als die jährliche Zunahme seit Beginn der Messungen im Jahr 1957.
Es wurde angenommen, dass diese Schwankungen auf die saisonale Aufnahme von CO zurückzuführen sind2 während der Sommermonate. Eine neu entwickelte Technik zur Bewertung des KKW des Ökosystems hat bestätigt, dass saisonale Schwankungen durch saisonale Veränderungen des CO bedingt sind2 Aufnahme durch Vegetation.[15][14] Dies hat viele Wissenschaftler und politische Entscheidungsträger dazu veranlasst, zu spekulieren, dass Ökosysteme verwaltet werden können, um Probleme mit der globalen Erwärmung zu lindern. Diese Art der Bewirtschaftung kann die Wiederaufforstung oder Änderung der Walderntepläne für viele Teile der Welt umfassen.
Zersetzung und Nährstoffkreislauf[edit]
Zersetzung und Nährstoffkreislauf sind für die Produktion von Ökosystembiomasse von grundlegender Bedeutung. Die meisten natürlichen Ökosysteme sind auf Stickstoff (N) begrenzt und die Biomasseproduktion ist eng mit dem N-Umsatz korreliert.[16][17]
Typischerweise ist der externe Nährstoffeintrag sehr gering und ein effizientes Recycling von Nährstoffen erhält die Produktivität.[5] Die Zersetzung von Pflanzenabfällen macht den größten Teil der Nährstoffe aus, die durch Ökosysteme recycelt werden (Abbildung 3). Die Zersetzungsraten von Pflanzenstreu hängen stark von der Streuqualität ab. Eine hohe Konzentration an Phenolverbindungen, insbesondere Lignin, in Pflanzenstreu wirkt sich verzögernd auf die Streuzersetzung aus.[18][19] Komplexere C-Verbindungen werden langsamer zersetzt und es kann viele Jahre dauern, bis sie vollständig abgebaut sind. Die Zersetzung wird typischerweise mit exponentiellem Zerfall beschrieben und steht im Zusammenhang mit den Mineralstoffkonzentrationen, insbesondere Mangan, in der Laubstreu.[20][21]


Weltweit werden die Zersetzungsraten durch die Abfallqualität und das Klima vermittelt.[22] Ökosysteme, die von Pflanzen mit niedriger Ligninkonzentration dominiert werden, weisen häufig schnelle Zersetzungs- und Nährstoffkreislaufraten auf (Chapin et al. 1982). Einfache Kohlenstoff (C) enthaltende Verbindungen werden bevorzugt durch Zersetzer-Mikroorganismen metabolisiert, was zu schnellen anfänglichen Zersetzungsraten führt, siehe 5A,[23] Modelle, die von konstanten Zerfallsraten abhängen; sogenannte “k” -Werte, siehe 5B.[24] Neben der Abfallqualität und dem Klima ist die Aktivität der Bodenfauna sehr wichtig [25]
Diese Modelle spiegeln jedoch nicht simultane lineare und nichtlineare Zerfallsprozesse wider, die wahrscheinlich während der Zersetzung auftreten. Zum Beispiel zersetzen sich Proteine, Zucker und Lipide exponentiell, aber Lignin zerfällt linearer[18] Somit wird der Abfallzerfall durch vereinfachende Modelle ungenau vorhergesagt.[26]
Ein einfaches alternatives Modell, das in 5C dargestellt ist, zeigt eine signifikant schnellere Zerlegung als das Standardmodell von 4B. Ein besseres Verständnis der Zersetzungsmodelle ist ein wichtiges Forschungsgebiet der Ökosystemökologie, da dieser Prozess eng mit der Nährstoffversorgung und der Gesamtkapazität der Ökosysteme zur Bindung von CO verbunden ist2 aus der Atmosphäre.
Trophäendynamik[edit]
Die trophische Dynamik bezieht sich auf den Prozess des Energie- und Nährstofftransfers zwischen Organismen. Die trophische Dynamik ist ein wichtiger Bestandteil der Struktur und Funktion von Ökosystemen. Abbildung 3 zeigt die Energieübertragung für ein Ökosystem in Silver Springs, Florida. Energie, die von Primärproduzenten (Pflanzen, P) gewonnen wird, wird von Pflanzenfressern (H) verbraucht, die von Fleischfressern (C) verbraucht werden, die selbst von „Top-Fleischfressern“ (TC) verbraucht werden.
Eines der offensichtlichsten Muster in Abbildung 3 ist, dass die Gesamtenergiemenge abnimmt, wenn man sich auf höhere trophische Ebenen bewegt (dh von Pflanzen zu Fleischfressern). Pflanzen üben eine „Bottom-up“ -Kontrolle auf die Energiestruktur von Ökosystemen aus, indem sie die Gesamtenergiemenge bestimmen, die in das System gelangt.[27]
Raubtiere können jedoch auch die Struktur niedrigerer trophischer Ebenen von oben nach unten beeinflussen. Diese Einflüsse können dominante Arten in terrestrischen und marinen Systemen dramatisch verändern[28][29] Das Zusammenspiel und die relative Stärke von Top-Down- und Bottom-Up-Kontrollen der Ökosystemstruktur und -funktion ist ein wichtiges Forschungsgebiet auf dem Gebiet der Ökologie.
Die trophische Dynamik kann die Zersetzungsraten und den Nährstoffkreislauf in Zeit und Raum stark beeinflussen. Zum Beispiel kann Pflanzenfresser die Zersetzung von Streu und den Nährstoffkreislauf durch direkte Änderungen der Streuqualität und eine veränderte dominante Vegetation erhöhen.[30] Es wurde gezeigt, dass Insektenfresser die Zersetzungsraten und den Nährstoffumsatz aufgrund von Änderungen der Streuqualität und erhöhten Frasseinträgen erhöhen.[1][31]
Der Ausbruch von Insekten erhöht jedoch nicht immer den Nährstoffkreislauf. Stadler[32] zeigten, dass C-reicher Honigtau, der während des Ausbruchs von Blattläusen produziert wird, zu einer erhöhten N-Immobilisierung durch Bodenmikroben führen kann, wodurch der Nährstoffkreislauf verlangsamt und möglicherweise die Biomasseproduktion eingeschränkt wird. Nordatlantische Meeresökosysteme wurden durch Überfischung von Kabeljau stark verändert. Kabeljaubestände stürzten in den 1990er Jahren ab, was zu einer Zunahme ihrer Beute wie Garnelen und Schneekrabben führte[29] Das Eingreifen des Menschen in Ökosysteme hat zu dramatischen Veränderungen der Struktur und Funktion des Ökosystems geführt. Diese Veränderungen treten rasch auf und haben unbekannte Folgen für die wirtschaftliche Sicherheit und das Wohlergehen der Menschen.[33]
Anwendungen und Bedeutung[edit]
Lehren aus zwei zentralamerikanischen Städten[edit]
Die Biosphäre wurde durch die Anforderungen der menschlichen Gesellschaften stark verändert. Die Ökosystemökologie spielt eine wichtige Rolle für das Verständnis und die Anpassung an die dringendsten aktuellen Umweltprobleme. Wiederherstellungsökologie und Ökosystemmanagement sind eng mit der Ökosystemökologie verbunden. Die Wiederherstellung stark verschlechterter Ressourcen hängt von der Integration der Funktionsmechanismen von Ökosystemen ab.[34]
Ohne diese intakten Funktionen wird der wirtschaftliche Wert von Ökosystemen stark verringert und es können sich potenziell gefährliche Bedingungen auf dem Gebiet entwickeln. Beispielsweise sind Gebiete im bergigen westlichen Hochland von Guatemala anfälliger für katastrophale Erdrutsche und lähmende saisonale Wasserknappheit aufgrund des Verlusts von Waldressourcen. Im Gegensatz dazu weisen Städte wie Totonicapán, die Wälder durch starke soziale Institutionen erhalten haben, eine größere lokale wirtschaftliche Stabilität und insgesamt ein größeres menschliches Wohlbefinden auf.[35]
Diese Situation ist bemerkenswert, wenn man bedenkt, dass diese Gebiete nahe beieinander liegen, die Mehrheit der Einwohner Maya-Abstammung hat und die Topographie und die Gesamtressourcen ähnlich sind. Dies ist ein Fall von zwei Personengruppen, die Ressourcen auf grundlegend unterschiedliche Weise verwalten. Die Ökosystemökologie liefert die Grundlagenforschung, die erforderlich ist, um eine Verschlechterung zu vermeiden und Ökosystemprozesse wiederherzustellen, die die Grundbedürfnisse des Menschen befriedigen.
Siehe auch[edit]
Verweise[edit]
- ^ ein b c Chapman, SK, Hart, SC, Cobb, NS, Whitham, TG und Koch, GW (2003). “Insektenfresser erhöhen die Streuqualität und -zersetzung: eine Erweiterung der Beschleunigungshypothese”. im: Ökologie 84: 2867 & ndash; 2876.
- ^ Hagen, JB (1992). Eine verschränkte Bank: Die Ursprünge der Ökosystemökologie. Rutgers University Press, New Brunswick, NJ
- ^ Odum, HT (1971). Umwelt, Macht und Gesellschaft. Wiley-Interscience New York, NY
- ^ Odum, EP 1969. “Die Strategie der Ökosystementwicklung”. im: Wissenschaft 164: 262 & ndash; 270.
- ^ ein b c Likens, GE, FH Bormann, NM Johnson, DW Fisher und RS Pierce. (1970). “Auswirkungen von Waldrodung und Herbizidbehandlung auf die Nährstoffbudgets im Ökosystem der Wasserscheide von Hubbard Brook”. im: Ökologische Monographien 40: 23-47.
- ^ Chapin, FS III, BH, Walker, RJ, Hobbs, DU, Hooper, JH, Lawton, OE, Sala und D., Tilman. (1997). “Biotische Kontrolle über das Funktionieren von Ökosystemen”. im: Wissenschaft 277: 500 & ndash; 504.
- ^ Defries, RS, JA Foley und GP Asner. (2004). “Landnutzungsentscheidungen: Gleichgewicht zwischen menschlichen Bedürfnissen und Ökosystemfunktion”. im: Grenzen in Ökologie und Umweltwissenschaften. 2: 249 & ndash; 257.
- ^ Chrispeels, MJ und Sadava, D. (1977). Pflanzen, Lebensmittel und Menschen. WH Freeman and Company, San Francisco.
- ^ Chinone, MA, NE Borlaug, CR Dowswell. (1997). “Eine auf Düngemitteln basierende grüne Revolution für Afrika”. Im: Wiederauffüllung der Bodenfruchtbarkeit in Afrika. Sonderpublikation der Soil Science Society of America Nr. 51. Soil Science Society of America, Madison, WI.
- ^ Foster, DR (1992). “Landnutzungsgeschichte (1730-1990) und Vegetationsdynamik in Zentral-New England, USA”. Im: Zeitschrift für Ökologie 80: 753 & ndash; 772.
- ^ Motzkin, G., DR Foster, A. Allen, J. Harrod und RD Boone. (1996). “Kontrollstelle zur Bewertung der Geschichte: Vegetationsmuster einer New England Sandfläche”. Im: Ökologische Monographien 66: 345 & ndash; 365.
- ^ Huxman TE, ea. (2004). “Konvergenz zwischen den Biomen zu einer gemeinsamen Effizienz der Regennutzung”. Natur. 429: 651 & ndash; 654
- ^ Keeling, CD und TP Whorf. (2005). Atmosphärisches CO2 Aufzeichnungen von Standorten im SIO-Luftprobenahmenetz “. In: Trends: Ein Kompendium von Daten zum globalen Wandel. Informationszentrum für Kohlendioxidinformation, Oak Ridge National Laboratory, US-Energieministerium, Oak Ridge, Tenn., USA
- ^ ein b Barford, CC, ea. (2001). Faktoren, die die langfristige und kurzfristige Sequestrierung von atmosphärischem CO steuern2 in einem Wald mittlerer Breite “. In: Wissenschaft 294: 1688 & ndash; 1691
- ^ Goulden, ML, JW Munger, S.-M. Fan, BC Daube und SC Wofsy (1996). “Auswirkungen der interannuellen Klimavariabilität auf den Kohlendioxidaustausch eines gemäßigten Laubwaldes”. Im: Wissenschaft 271: 1576 & ndash; 1578
- ^ Vitousek, PM und Howarth, RW (1991). “Stickstoffbegrenzung an Land und im Meer: Wie kann sie auftreten?” Im: Biogeochemie 13: 87-115.
- ^ Reich, PB, Grigal, DF, Aber, JD, Gower, ST (1997). “Stickstoffmineralisierung und Produktivität in 50 Hartholz- und Nadelbäumen stehen auf verschiedenen Böden”. Im: Ökologie 78: 335 & ndash; 347.
- ^ ein b Melillo, JM, Aber, JD und Muratore, JF (1982). “Stickstoff- und Ligninkontrolle der Zersetzungsdynamik von Hartholzblattstreu”. Im: Ökologie 63: 621 & ndash; 626.
- ^ Hättenschwiler S. und PM Vitousek (2000). “Die Rolle von Polyphenolen im Nährstoffkreislauf des terrestrischen Ökosystems”. Im: Trends in Ökologie und Evolution 15: 238-243
- ^ Davey MP, B. Berg, P. Rowland, BA Emmett. 2007. Die Zersetzung von Eichenblattstreu hängt mit den anfänglichen Mn-Konzentrationen des Streus zusammen. Kanadisches Journal für Botanik. 85 (1). 16-24.
- ^ Berg B., Davey MP, Emmett B., Faituri M., Hobbie S., Johansson MB, Liu C., De Marco A., McClaugherty C., Norell L., Rutigliano F., De Santo AV. 2010. Einflussfaktoren auf Grenzwerte für die Zersetzung von Kiefernnadelstreu – eine Synthese für boreale und gemäßigte Kiefernwaldsysteme. Biogeochemie. 100: 57 & ndash; 73
- ^ Meentemeyer, V. 1978 “Makroklima- und Ligninkontrolle der Streuzersetzungsraten”. im: Ökologie 59: 465 & ndash; 472.
- ^ Aber, JD und JM, Melillo (1982). “Stickstoffimmobilisierung in verrottendem Hartholzblattstreu als Funktion des anfänglichen Stickstoff- und Ligningehalts”. Im: Kanadisches Journal für Botanik 60: 2263 & ndash; 2269.
- ^ Olson, JS (1963). “Energiespeicherung und das Gleichgewicht von Produzenten und Zersetzern in ökologischen Systemen”. Im: Ökologie 44: 322 & ndash; 331.
- ^ Castro-Huerta, R.; Falco, L.; Sandler, R.; Coviella, C. (2015). “Unterschiedlicher Beitrag von Boden-Biota-Gruppen zur Zersetzung von Pflanzenstreu, vermittelt durch Bodennutzung”. PeerJ. 3: e826. doi:10.7717 / peerj.826. PMC 4359044. PMID 25780777.
- ^ Carpenter, SA (1981). “Zerfall von heterogenem Detritus: ein allgemeines Modell”. Im: Zeitschrift für theoretische Biologie 89: 539 & ndash; 547.
- ^ Chapin FS III, Matson, PA und Mooney, HA (2003). Prinzipien der terrestrischen Ökosystemökologie. Springer-Verlag, New York, NY
- ^ Belovsky, GE und JB Slade. (2000). “Insektenfresser beschleunigen den Nährstoffkreislauf und steigern die Pflanzenproduktion”. Im: Verfahren der Nationalen Akademie der Wissenschaften (VEREINIGTE STAATEN VON AMERIKA). 97: 14412-14417.
- ^ ein b Frank et al. 2005.
- ^ Hunter, MD (2001). “Die Dynamik der Insektenpopulation trifft auf die Ökosystemökologie: Auswirkungen von Pflanzenfressern auf die Nährstoffdynamik des Bodens”. Im: Land- und Forstentomologie 3: 77-84.
- ^ Swank, WT, Waide, JB, Crossley, DA und Todd RL (1981). “Die Entlaubung von Insekten verbessert den Nitrat-Export aus Waldökosystemen”. Im: Oecologia 51: 297 & ndash; 299.
- ^ Stadler, B., Solinger, St. und Michalzik, B. (2001). “Insektenfresser und der Nährstofffluss vom Baldachin zum Boden in Nadel- und Laubwäldern”. Im: Oecologia 126: 104 & ndash; 113
- ^ Die enge Beziehung zwischen Ökosystemleistungen und Wohlbefinden wurde in der Millennium Ecosystem Assessment hervorgehoben
- ^ Ehrenfeld, JG und Toth, LA (1997). “Restaurierungsökologie und Ökosystemperspektive”. im: Restaurierungsökologie 5: 307-317.
- ^ Conz, BW 2004. Kontinuität und Anfechtung: Naturschutzlandschaften in Totonicapán, Guatemala. Masterarbeit der University of Massachusetts.
Recent Comments