Hexokinase – Wikipedia
EIN Hexokinase ist ein Enzym, das Hexosen (Zucker mit sechs Kohlenstoffatomen) phosphoryliert und Hexosephosphat bildet. In den meisten Organismen ist Glucose das wichtigste Substrat für Hexokinasen, und Glucose-6-phosphat ist das wichtigste Produkt. Hexokinase besitzt die Fähigkeit, eine anorganische Phosphatgruppe von ATP auf ein Substrat zu übertragen.
Hexokinasen sollten nicht mit Glucokinase verwechselt werden, einer spezifischen Isoform der Hexokinase. Alle Hexokinasen sind in der Lage, mehrere Hexosen zu phosphorylieren, aber Glucokinase wirkt mit einer 50-fach niedrigeren Substrataffinität und sein Haupthexosesubstrat ist Glucose.
Variation[edit]
Gene, die für Hexokinase kodieren, wurden in allen Lebensbereichen entdeckt und existieren unter einer Vielzahl von Arten, die von Bakterien, Hefen und Pflanzen bis hin zu Menschen und anderen Wirbeltieren reichen. Sie sind kategorisiert als Aktinfalte Proteine, die einen gemeinsamen ATP-Bindungsstellenkern teilen, der von variableren Sequenzen umgeben ist, die Substrataffinitäten und andere Eigenschaften bestimmen.
In einer Spezies können mehrere Hexokinase-Isoformen oder Isozyme auftreten, die unterschiedliche Funktionen bieten.
Reaktion[edit]
Die durch Hexokinasen vermittelten intrazellulären Reaktionen können wie folgt charakterisiert werden:
- Hexose-CH2OH + MgATP2−
→ Hexose-CH2O-PO2−
3 + MgADP– –
+ H.+
wo Hexose-CH2OH repräsentiert eine von mehreren Hexosen (wie Glucose), die ein zugängliches -CH enthalten2OH-Einheit.
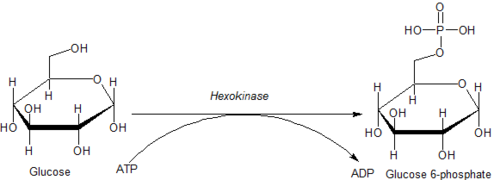
Folgen der Hexosephosphorylierung[edit]
Die Phosphorylierung einer Hexose wie Glucose beschränkt sie häufig auf eine Reihe von intrazellulären Stoffwechselprozessen wie Glykolyse oder Glykogensynthese. Dies liegt daran, dass phosphorylierte Hexosen geladen sind und daher schwieriger aus einer Zelle zu transportieren sind.
Bei Patienten mit essentieller Fructosurie ist der Metabolismus von Fructose durch Hexokinase zu Fructose-6-phosphat die primäre Methode zur Metabolisierung von Fructose aus der Nahrung. Dieser Weg ist bei normalen Personen nicht signifikant.
Größe verschiedener Isoformen[edit]
Die meisten bakteriellen Hexokinasen sind ungefähr 50 kD groß. Mehrzellige Organismen, einschließlich Pflanzen und Tiere, weisen häufig mehr als eine Hexokinase-Isoform auf. Die meisten sind etwa 100 kD groß und bestehen aus zwei Hälften (N- und C-Terminal), die viel Sequenzhomologie gemeinsam haben. Dies deutet auf einen evolutionären Ursprung durch Duplikation und Fusion einer 50 kD-Hexokinase hin, die denen von Bakterien ähnlich ist.
Arten von Säugetierhexokinase[edit]
Es gibt vier wichtige Hexokinase-Isozyme von Säugetieren (EC 2.7.1.1), die sich in subzellulären Positionen und Kinetiken in Bezug auf verschiedene Substrate und Bedingungen sowie die physiologische Funktion unterscheiden. Sie werden als Hexokinasen I, II, III und IV oder Hexokinasen A, B, C und D bezeichnet.
Hexokinasen I, II und III[edit]
Die Hexokinasen I, II und III werden als “Low-K” bezeichnetmIsozyme aufgrund einer hohen Affinität für Glucose (unter 1 mM). Hexokinasen I und II folgen der Michaelis-Menten-Kinetik bei physiologischen Konzentrationen von Substraten.[citation needed] Alle drei werden durch ihr Produkt Glucose-6-phosphat stark gehemmt. Molekulargewichte liegen bei ca. 100 kD. Jede besteht aus zwei ähnlichen 50-kD-Hälften, aber nur in Hexokinase II haben beide Hälften funktionelle aktive Stellen.
- Hexokinase I / A kommt in allen Geweben von Säugetieren vor und wird als “Haushaltsenzym” angesehen, das von den meisten physiologischen, hormonellen und metabolischen Veränderungen nicht beeinflusst wird.
- Hexokinase II / B stellt bei vielen Zelltypen die hauptsächliche regulierte Isoform dar und ist bei vielen Krebsarten erhöht. Es ist die Hexokinase, die in Muskel und Herz gefunden wird. Hexokinase II befindet sich auch an der äußeren Membran der Mitochondrien, so dass sie direkten Zugang zu ATP haben kann.[2] Die relative spezifische Aktivität von Hexokinase II steigt mit dem pH-Wert mindestens in einem pH-Bereich von 6,9 bis 8,5 an.[3]
- Hexokinase III / C wird in physiologischen Konzentrationen durch Glucose substrathemmt. Über die regulatorischen Eigenschaften dieser Isoform ist wenig bekannt.
Hexokinase IV (“Glucokinase”)[edit]
Die Hexokinase IV von Säugetieren, auch als Glucokinase bezeichnet, unterscheidet sich von anderen Hexokinasen in Kinetik und Funktionen.
Der Ort der Phosphorylierung auf subzellulärer Ebene tritt auf, wenn Glucokinase zwischen dem Zytoplasma und dem Zellkern von Leberzellen transloziert. Glucokinase kann Glucose nur phosphorylieren, wenn die Konzentration dieses Substrats hoch genug ist; sein Km für Glucose ist 100-mal höher als der der Hexokinasen I, II und III.
Hexokinase IV ist monomer, etwa 50 kD, zeigt eine positive Kooperativität mit Glucose und wird durch sein Produkt Glucose-6-phosphat nicht allosterisch gehemmt.
Hexokinase IV ist in Leber, Bauchspeicheldrüse, Hypothalamus, Dünndarm und möglicherweise bestimmten anderen neuroendokrinen Zellen vorhanden und spielt eine wichtige regulatorische Rolle im Kohlenhydratstoffwechsel. In den Beta-Zellen der Pankreasinseln dient es als Glukosesensor zur Steuerung der Insulinfreisetzung und steuert in ähnlicher Weise die Glukagonfreisetzung in den Alpha-Zellen. In Hepatozyten der Leber reagiert die Glukokinase auf Änderungen der Umgebungsglukosespiegel, indem sie die Glykogensynthese erhöht oder verringert.
In der Glykolyse[edit]
Glucose ist insofern einzigartig, als sie von allen Zellen sowohl in Gegenwart als auch in Abwesenheit von molekularem Sauerstoff (O) zur Herstellung von ATP verwendet werden kann2). Der erste Schritt bei der Glykolyse ist die Phosphorylierung von Glucose durch Hexokinase.
Verbindung C00031 bei KEGG Pathway Database. Enzym 2.7.1.1 bei KEGG Pathway Database. Verbindung C00668 bei KEGG Pathway Database. Reaktion R01786 bei KEGG Pathway Database.
Durch die Katalyse der Phosphorylierung von Glucose zu Glucose-6-phosphat behalten Hexokinasen den Konzentrationsgradienten bei, der den erleichterten Transport von Glucose in Zellen begünstigt. Diese Reaktion initiiert auch alle physiologisch relevanten Wege der Glukoseverwertung, einschließlich der Glykolyse und des Pentosephosphatweges.[4] Die Zugabe einer geladenen Phosphatgruppe an der 6-Position von Hexosen stellt auch das “Einfangen” von Glucose- und 2-Desoxyhexose-Glucoseanaloga (z. B. 2-Desoxyglucose und 2-Fluor-2-desoxyglucose) in Zellen sicher, wie es geladene Hexosephosphate nicht können leicht die Zellmembran überqueren.
Assoziation mit Mitochondrien[edit]
Die Hexokinasen I und II können sich durch spezifische Bindung an ein Porin oder einen spannungsabhängigen Anionenkanal physikalisch mit der Außenfläche der äußeren Membran der Mitochondrien verbinden. Diese Assoziation verleiht der Hexokinase direkten Zugang zu ATP, das von Mitochondrien erzeugt wird, einem der beiden Substrate der Hexokinase. Mitochondriale Hexokinase ist in schnell wachsenden malignen Tumorzellen stark erhöht und weist bis zu 200-mal höhere Werte als normales Gewebe auf. Es wurde gezeigt, dass mitochondrial gebundene Hexokinase die treibende Kraft ist[5] für die extrem hohen glykolytischen Raten, die aerob in Tumorzellen stattfinden (der sogenannte Warburg-Effekt, der 1930 von Otto Heinrich Warburg beschrieben wurde).
Hydropathie-Handlung[edit]

Die potentiellen Transmembranteile eines Proteins können durch Hydropathieanalyse nachgewiesen werden. Eine Hydropathieanalyse verwendet einen Algorithmus, der den hydrophoben Charakter an jeder Position entlang der Polypeptidkette quantifiziert. Eine der akzeptierten Hydropathie-Skalen ist die von Kyte und Doolittle, die auf der Erzeugung von Hydropathie-Plots beruht. In diesen Darstellungen repräsentieren die negativen Zahlen hydrophile Regionen und die positiven Zahlen hydrophobe Regionen auf der y-Achse. Eine potentielle Transmembrandomäne ist auf der x-Achse etwa 20 Aminosäuren lang.
Nach diesen Standards wurde eine Hydropathieanalyse der Hexokinase in Hefe erstellt. Es scheint, als ob Hexokinase eine einzige potentielle Transmembrandomäne besitzt, die sich um Aminosäure 400 befindet. Daher ist Hexokinase höchstwahrscheinlich kein integrales Membranprotein in Hefe.[6]
Mangel[edit]
Hexokinase-Mangel ist eine genetisch autosomal-rezessive Erkrankung, die eine chronische hämolytische Anämie verursacht. Chronische hämolytische Anämie wird durch eine Mutation im HK-Gen verursacht, das für das HK-Enzym kodiert. Die Mutation bewirkt eine Verringerung der HK-Aktivität, was zu einem Hexokinasemangel führt.[7]
Siehe auch[edit]
Verweise[edit]
- ^ PDB: 308;; Kuettner EB, Kettner K, Keim A, Svergun DI, Volke D (2010). “Kristallstruktur von dimerem KlHxk1 in Kristallform I”. doi:10.2210 / pdb3o08 / pdb.
- ^ “Hexokinase-Daten zu Uniprot”. uniprot.org.
- ^ Šimčíková D, Heneberg P (August 2019). “Identifizierung des alkalischen pH-Optimums der menschlichen Glucokinase aufgrund der ATP-vermittelten Bias-Korrektur in den Ergebnissen von Enzymtests”. Wissenschaftliche Berichte. 9 (1): 11422. doi:10.1038 / s41598-019-47883-1. PMC 6684659. PMID 31388064.
- ^ Robey, RB; Hay, N. (2006). “Mitochondriale Hexokinasen, neuartige Mediatoren der antiapoptotischen Wirkung von Wachstumsfaktoren und Akt”. Onkogen. 25 (34): 4683–96. doi:10.1038 / sj.onc.1209595. PMID 16892082.
- ^ Bustamante E, Pedersen P (1977). “Hohe aerobe Glykolyse von Rattenhepatomzellen in Kultur: Rolle der mitochondrialen Hexokinase”. Proc Natl Acad Sci USA. 74 (9): 3735–9. Bibcode:1977PNAS … 74.3735B. doi:10.1073 / pnas.74.9.3735. PMC 431708. PMID 198801.
- ^ Bowen, RA Molecular Toolkit: Proteinhydrophobizitätsdiagramme. Colorado State University, 1998. Web. 15. November 2010.http://www.vivo.colostate.edu/molkit/index.html Archiviert 25. Juni 2010 an der Wayback-Maschine>
- ^ “Hexokinase-Mangel”. Enerca. Enerca. Abgerufen 6. April 2017.
Recent Comments