Herkunft der Vögel – Wikipedia


Die wissenschaftliche Frage, innerhalb derer sich eine größere Gruppe von Tiervögeln entwickelte, wurde traditionell als “Herkunft der Vögel‘. Der gegenwärtige wissenschaftliche Konsens ist, dass Vögel eine Gruppe von Maniraptoran-Theropod-Dinosauriern sind, die während des Mesozoikums entstanden sind.
Eine enge Beziehung zwischen Vögeln und Dinosauriern wurde erstmals im 19. Jahrhundert nach der Entdeckung des primitiven Vogels vorgeschlagen Archaeopteryx in Deutschland. Vögel und ausgestorbene Nicht-Vogel-Dinosaurier teilen viele einzigartige Skelettmerkmale.[1] Darüber hinaus wurden Fossilien von mehr als 30 Arten von Nicht-Vogel-Dinosauriern mit erhaltenen Federn gesammelt. Es gibt sogar sehr kleine Dinosaurier wie Microraptor und Anchiornis, die lange, verschwundene Arm- und Beinfedern haben, die Flügel bilden. Der jurassische basale Avialan Pedopenna zeigt auch diese langen Fußfedern. Der Paläontologe Lawrence Witmer kam 2009 zu dem Schluss, dass diese Beweise ausreichen, um zu zeigen, dass die Vogelentwicklung ein vierflügeliges Stadium durchlaufen hat.[2] Fossile Beweise zeigen auch, dass Vögel und Dinosaurier Merkmale wie hohle, pneumatisierte Knochen, Gastrolithen im Verdauungssystem, Nestbau und Brutverhalten gemeinsam hatten.
Obwohl der Ursprung von Vögeln historisch ein umstrittenes Thema in der Evolutionsbiologie war, bestreiten nur noch wenige Wissenschaftler den dinosaurischen Ursprung von Vögeln, was auf eine Abstammung von anderen Arten von archosaurischen Reptilien hindeutet. Innerhalb des Konsenses, der die Abstammung der Dinosaurier unterstützt, ist die genaue Abfolge der evolutionären Ereignisse, die die frühen Vögel innerhalb der Maniraptoran-Theropoden hervorgebracht haben, umstritten. Der Ursprung des Vogelfluges ist eine separate, aber verwandte Frage, für die auch mehrere Antworten vorgeschlagen werden.
Forschungsgeschichte[edit]
Huxley, Archaeopteryx und frühe Forschung[edit]


Die wissenschaftliche Untersuchung der Herkunft von Vögeln begann kurz nach der Veröffentlichung von Charles Darwins im Jahr 1859 Über den Ursprung der Arten.[3] 1860 wurde im deutschen spätnurassischen Solnhofen-Kalkstein eine versteinerte Feder entdeckt. Christian Erich Hermann von Meyer beschrieb diese Feder als Archaeopteryx lithographica Das nächste Jahr.[4]Richard Owen beschrieb 1863 ein fast vollständiges Skelett und erkannte es trotz vieler Merkmale, die an Reptilien erinnern, einschließlich Krallenvorderbeinen und eines langen, knöchernen Schwanzes, als Vogel an.[5]
Der Biologe Thomas Henry Huxley, bekannt als “Darwins Bulldogge” für seine hartnäckige Unterstützung der neuen Evolutionstheorie durch natürliche Selektion, wurde fast sofort aufgegriffen Archaeopteryx als Übergangsfossil zwischen Vögeln und Reptilien. Ab 1868 und nach früheren Vorschlägen von Karl Gegenbaur,[6] und Edward Drinker Cope,[7] Huxley machte detaillierte Vergleiche von Archaeopteryx mit verschiedenen prähistorischen Reptilien und stellte fest, dass es Dinosauriern am ähnlichsten war Hypsilophodon und Compsognathus.[8][9] Die Entdeckung des legendären “Berliner Exemplars” von Archaeopteryx, komplett mit einem Satz Reptilienzähne, lieferte weitere Beweise. Wie Cope schlug Huxley eine evolutionäre Beziehung zwischen Vögeln und Dinosauriern vor. Obwohl Huxley von dem sehr einflussreichen Owen abgelehnt wurde, wurden seine Schlussfolgerungen von vielen Biologen akzeptiert, darunter Baron Franz Nopcsa.[10] während andere, insbesondere Harry Seeley,[11] argumentierte, dass die Ähnlichkeiten auf konvergente Evolution zurückzuführen seien.
Heilmann und die Thecodont-Hypothese[edit]
Ein Wendepunkt kam im frühen zwanzigsten Jahrhundert mit den Schriften von Gerhard Heilmann aus Dänemark. Heilmann war von Beruf Künstler und interessierte sich wissenschaftlich für Vögel. Von 1913 bis 1916 erweiterte er frühere Arbeiten von Othenio Abel.[12] veröffentlichte die Ergebnisse seiner Forschung in mehreren Teilen, die sich mit Anatomie, Embryologie, Verhalten, Paläontologie und Evolution von Vögeln befassten.[13] Seine Arbeit, ursprünglich in dänischer Sprache geschrieben als Vor Nuvaerende Viden om Fuglenes Afstamning, wurde zusammengestellt, ins Englische übersetzt und 1926 als veröffentlicht Der Ursprung der Vögel.


Wie Huxley verglich Heilmann Archaeopteryx und andere Vögel zu einer erschöpfenden Liste prähistorischer Reptilien und kamen auch zu dem Schluss, dass Theropodendinosaurier mögen Compsognathus waren am ähnlichsten. Heilmann stellte jedoch fest, dass Vögel Schlüsselbeine (Schlüsselbeine) hatten, die zu einem Knochen namens Furcula (“Querlenker”) verschmolzen waren, und obwohl Schlüsselbeine bei primitiveren Reptilien bekannt waren, waren sie bei Dinosauriern noch nicht erkannt worden. Da er fest an Dollos Gesetz glaubte, das besagt, dass die Evolution nicht reversibel ist, konnte Heilmann nicht akzeptieren, dass Schlüsselbeine bei Dinosauriern verloren gingen und sich bei Vögeln neu entwickelten. Er war daher gezwungen, Dinosaurier als Vogelvorfahren auszuschließen und all ihre Ähnlichkeiten der konvergenten Evolution zuzuschreiben. Heilmann erklärte, dass Vogelvorfahren stattdessen unter den primitiveren “Thecodont” -Reptilien gefunden würden.[14] Heilmanns äußerst gründlicher Ansatz sorgte dafür, dass sein Buch zu einem Klassiker auf diesem Gebiet wurde und seine Schlussfolgerungen zur Vogelherkunft, wie bei den meisten anderen Themen, von fast allen Evolutionsbiologen für die nächsten vier Jahrzehnte akzeptiert wurden.[15]
Schlüsselbeine sind relativ empfindliche Knochen und daher in Gefahr, bis zur Unkenntlichkeit zerstört oder zumindest beschädigt zu werden. Trotzdem waren einige fossile Theropoden-Schlüsselbeine tatsächlich ausgegraben worden, bevor Heilmann sein Buch schrieb, aber diese waren falsch identifiziert worden.[16]
Das Fehlen von Schlüsselbeinen bei Dinosauriern wurde trotz der Entdeckung von Schlüsselbeinen im primitiven Theropoden zur orthodoxen Ansicht Segisaurus im Jahr 1936.[17] Der nächste Bericht über Schlüsselbeine bei einem Dinosaurier war 1983 in einem russischen Artikel.[18]
Entgegen der Überzeugung Heilmanns akzeptieren Paläontologen heute, dass Schlüsselbeine und in den meisten Fällen Furculae nicht nur bei Theropoden, sondern auch bei saurischen Dinosauriern Standard sind. Bis Ende 2007 wurden verknöcherte Furculae (dh eher aus Knochen als aus Knorpel) in allen Arten von Theropoden gefunden, mit Ausnahme der basalsten. Eoraptor und Herrerasaurus.[19] Der ursprüngliche Bericht einer Furcula im primitiven Theropoden Segisaurus (1936) wurde durch eine erneute Untersuchung im Jahr 2005 bestätigt.[20] Verbundene, furculaähnliche Schlüsselbeine wurden ebenfalls in gefunden Massospondylus, ein frühes Jura-Sauropodomorph.[21]
Ostrom, Deinonychus und die Renaissance der Dinosaurier[edit]


Nach der Entdeckung eines neuen Theropodendinosauriers in Montana im Jahr 1964 begann sich das Blatt gegen die Hypothese des „Thecodont“ zu wenden. 1969 wurde dieser Dinosaurier beschrieben und benannt Deinonychus von John Ostrom von der Yale University.[22] Im nächsten Jahr beschrieb Ostrom ein Exemplar von neu Pterodactylus im niederländischen Teyler Museum als weiteres Skelett von Archaeopteryx.[23] Das Exemplar bestand hauptsächlich aus einem einzigen Flügel und seine Beschreibung machte Ostrom auf die Ähnlichkeiten zwischen den Handgelenken von aufmerksam Archaeopteryx und Deinonychus.[24]
1972 stellte der britische Paläontologe Alick Walker die Hypothese auf, dass Vögel nicht aus “Thecodonten”, sondern aus Krokodil-Vorfahren wie stammten Sphenosuchus.[25] Ostroms Arbeit mit Theropoden und Frühaufstehern veranlasste ihn, Mitte der 1970er Jahre mit einer Reihe von Veröffentlichungen zu antworten, in denen er die vielen Ähnlichkeiten zwischen Vögeln und Theropodendinosauriern darlegte und die Ideen wiederbelebte, die Huxley vor über einem Jahrhundert erstmals vorgebracht hatte.[26][27][28] Ostroms Anerkennung der dinosaurischen Abstammung von Vögeln, zusammen mit anderen neuen Ideen über den Stoffwechsel von Dinosauriern,[29] Aktivitätsniveaus und elterliche Fürsorge,[30] begann die sogenannte Dinosaurier-Renaissance, die in den 1970er Jahren begann und bis heute andauert.
Ostroms Enthüllungen fielen auch mit der zunehmenden Übernahme der phylogenetischen Systematik (Kladistik) zusammen, die in den 1960er Jahren mit der Arbeit von Willi Hennig begann.[31] Die Kladistik ist eine exakte Methode zur Anordnung von Arten, die ausschließlich auf ihren evolutionären Beziehungen basiert. Diese werden berechnet, indem der Evolutionsbaum bestimmt wird, der die geringste Anzahl von Änderungen ihrer anatomischen Eigenschaften impliziert. In den 1980er Jahren wurde die kladistische Methodik erstmals von Jacques Gauthier und anderen auf die Dinosaurierphylogenie angewendet, was eindeutig zeigt, dass Vögel eine abgeleitete Gruppe von Theropodendinosauriern waren.[32] Frühe Analysen deuteten darauf hin, dass Dromaeosaurid-Theropoden mögen Deinonychus waren besonders eng mit Vögeln verwandt, ein Ergebnis, das seitdem vielfach bestätigt wurde.[33][34]
Gefiederte Dinosaurier in China[edit]


In den frühen neunziger Jahren wurden spektakulär erhaltene Vogelfossilien in mehreren geologischen Formationen der frühen Kreidezeit in der nordöstlichen chinesischen Provinz Liaoning entdeckt.[35][36] 1996 beschrieben chinesische Paläontologen Sinosauropteryx als neue Vogelgattung aus der Yixian-Formation,[37] Aber dieses Tier wurde schnell als basaler Theropodendinosaurier erkannt, der eng mit ihm verwandt ist Compsognathus. Überraschenderweise war sein Körper von langen filamentösen Strukturen bedeckt. Diese wurden als “Protofedern” bezeichnet und als homolog mit den fortgeschritteneren Vogelfedern angesehen.[38] obwohl einige Wissenschaftler mit dieser Einschätzung nicht einverstanden sind.[39] Chinesische und nordamerikanische Wissenschaftler beschrieben Caudipteryx und Protarchaeopteryx bald darauf. Aufgrund der Skelettmerkmale waren diese Tiere Nicht-Vogel-Dinosaurier, aber ihre Überreste trugen vollständig geformte Federn, die denen von Vögeln sehr ähnlich waren.[40] “Archaeoraptor”, beschrieben ohne Peer Review in einer Ausgabe von 1999 von National Geographic,[41] stellte sich als geschmuggelte Fälschung heraus,[42] Aber legitime Überreste strömen weiterhin aus dem Yixian, sowohl legal als auch illegal. Federn oder “Protofedern” wurden auf einer Vielzahl von Theropoden im Yixian gefunden.[43][44] und die Entdeckungen extrem vogelähnlicher Nicht-Vogel-Dinosaurier,[45] sowie nicht-aviärdinosaurierähnliche primitive Vögel,[46] haben die morphologische Lücke zwischen Nicht-Vogel-Theropoden und Vögeln fast vollständig geschlossen.
Ziffernhomologie[edit]
Es gibt eine Debatte zwischen Embryologen und Paläontologen, ob die Hände von Theropodendinosauriern und Vögeln wesentlich unterschiedlich sind, basierend auf der Phalangealzahl, einer Zählung der Anzahl der Phalangen (Finger) in der Hand. Dies ist ein wichtiges und heftig diskutiertes Forschungsgebiet, da seine Ergebnisse den Konsens in Frage stellen können, dass Vögel (Nachkommen von) Dinosauriern sind.
Embryologen und einige Paläontologen, die sich der Vogel-Dinosaurier-Verbindung widersetzen, haben die Ziffern der Vögel II-III-IV auf der Grundlage mehrerer Studien zur Entwicklung des Eies lange nummeriert.[47] Dies basiert auf der Tatsache, dass bei den meisten Amnioten die erste Ziffer, die sich in einer 5-Finger-Hand bildet, die Ziffer IV ist, die eine Primärachse entwickelt. Daher haben Embryologen die Primärachse bei Vögeln als Ziffer IV und die überlebenden Ziffern als II-III-IV identifiziert. Die Fossilien der Hände fortgeschrittener Theropoden (Tetanurae) scheinen die Ziffern I-II-III zu haben (einige Gattungen innerhalb von Avetheropoda haben auch eine reduzierte Ziffer IV[48]). Wenn dies zutrifft, ist die II-III-IV-Entwicklung von Ziffern bei Vögeln ein Hinweis auf die Abstammung von Theropoden (Dinosauriern). Da es jedoch keine ontogene (Entwicklungs-) Grundlage gibt, um definitiv anzugeben, welche Ziffern welche auf einer Theropodenhand sind (da heute kein Wachstum und keine Entwicklung von Nicht-Vogel-Theropoden beobachtet werden kann), ist die Kennzeichnung der Theropodenhand nicht absolut schlüssig.
Paläontologen haben Vogelziffern traditionell als I-II-III identifiziert. Sie argumentieren, dass die Ziffern der Vögel die Nummern I-II-III haben, genau wie die der Theropodendinosaurier, nach der konservierten Phalangealformel. Die Phalangealzahl für Archosaurier beträgt 2-3-4-5-3; Viele Archosaurier-Linien haben eine reduzierte Anzahl von Ziffern, aber die gleiche Phalangealformel in den verbleibenden Ziffern. Mit anderen Worten, Paläontologen behaupten, dass Archosaurier verschiedener Abstammungslinien dazu neigen, die gleichen Ziffern zu verlieren, wenn ein Ziffernverlust von außen nach innen auftritt. Die drei Ziffern der Dromaeosaurier und Archaeopteryx haben die gleiche Phalangealformel von I-II-III wie die Ziffern I-II-III von basalen Archosauriern. Daher wären die verlorenen Ziffern V und IV. Wenn dies wahr ist, würden moderne Vögel auch die Ziffern I-II-III besitzen.[47]
In einer Veröffentlichung aus dem Jahr 1999 wurde außerdem eine Frame-Verschiebung der Ziffern der Theropodenlinie vorgeschlagen, die zu Vögeln führt (wodurch Ziffer I zu Ziffer II, II zu III usw. wird).[49][50]
Solche Rahmenverschiebungen sind jedoch bei Amnioten selten und müssten – um mit dem Theropodenursprung der Vögel übereinzustimmen – nur in den Vorderbeinen der Vogel-Theropoden-Linie und nicht in den Hinterbeinen auftreten (ein Zustand, der bei keinem Tier unbekannt ist).[51]
Das nennt man Seitliche Ziffernreduzierung (LDR) versus Bilaterale Ziffernreduzierung (BDR) (siehe auch Limusaurus[52]
Eine kleine Minderheit, bekannt unter dem Akronym BAND (Vögel sind keine Dinosaurier)[53] darunter die Ornithologen Alan Feduccia und Larry Martin behaupten weiterhin, dass Vögel enger mit früheren Reptilien verwandt sind, wie z Longisquama oder Euparkeriaals zu Dinosauriern.[54][55]Embryologische Studien zur Vogelentwicklungsbiologie haben Fragen zur Ziffernhomologie in den Vorderbeinen von Vögeln und Dinosauriern aufgeworfen.[56] Aufgrund der überzeugenden Beweise aus der vergleichenden Anatomie und Phylogenetik sowie der dramatischen Fossilien von gefiederten Dinosauriern aus China findet die Idee, dass Vögel Dinosaurier sind, die zuerst von Huxley und später von Nopcsa und Ostrom verfochten wurden, nahezu einstimmige Unterstützung heutige Paläontologen.[15]
Thermogene Muskelhypothese[edit]
Eine Veröffentlichung aus dem Jahr 2011 schlug vor, dass die Auswahl für die Erweiterung des Skelettmuskels und nicht für die Entwicklung des Fluges die treibende Kraft für die Entstehung dieser Gruppe war.[57][58] Nach dieser Hypothese wurden die Muskeln bei prospektiv endothermen Sauriern größer als Reaktion auf den Verlust des mitochondrialen Entkopplungsproteins der Wirbeltiere, UCP1,[59] das ist thermogen. Bei Säugetieren wirkt UCP1 im braunen Fettgewebe, um Neugeborene vor Unterkühlung zu schützen. Bei modernen Vögeln hat der Skelettmuskel eine ähnliche Funktion und es wird vermutet, dass dies bei ihren Vorfahren der Fall war. Aus dieser Sicht waren Bipedalität und andere Veränderungen des Vogelgerüsts Nebenwirkungen der Muskelhyperplasie, wobei weitere evolutionäre Modifikationen der Vorderbeine, einschließlich Anpassungen für Flug oder Schwimmen, und Reste sekundäre Folgen der Zweibeinigkeit waren.
Phylogenie[edit]
Archaeopteryx wurde historisch als der erste Vogel angesehen, oder Urvogel. Obwohl neuere Fossilienfunde die Lücke zwischen Theropoden und Theropoden füllten Archaeopteryxsowie die Lücke zwischen Archaeopteryx und moderne Vögel, phylogenetische Taxonomen, verwenden im Einklang mit der Tradition fast immer Archaeopteryx als Spezifizierer zur Definition von Aves.[60][61] Aves wurde seltener als eine Kronengruppe definiert, die nur aus modernen Vögeln besteht.[32] Fast alle Paläontologen betrachten Vögel als coelurosaurische Theropodendinosaurier.[15] Innerhalb von Coelurosauria haben mehrere kladistische Analysen Unterstützung für eine Gruppe namens Maniraptora gefunden, die aus Therizinosauroiden, Oviraptorosauriern, Troodontiden, Dromaeosauriden und Vögeln besteht.[33][34][62] Von diesen sind Dromaeosauriden und Troodontiden normalerweise in der Klade Deinonychosauria vereint, die eine Schwestergruppe von Vögeln (die zusammen die Knotenklade Eumaniraptora bilden) innerhalb der Stammkladenparaven ist.[33][63]
Andere Studien haben alternative Phylogenien vorgeschlagen, bei denen sich bestimmte Gruppen von Dinosauriern, die normalerweise als Nicht-Vögel gelten, möglicherweise von Vorfahren der Vögel entwickelt haben. Eine Analyse aus dem Jahr 2002 ergab beispielsweise, dass Oviraptorosaurier basale Vögel waren.[64]Alvarezsauriden, bekannt aus Asien und Amerika, wurden verschiedentlich als basale Maniraptorane klassifiziert.[33][34][65][66] Paravianer,[62] das Schwestertaxon der Ornithomimosaurier,[67] sowie spezialisierte Frühaufsteher.[68][69] Die Gattung Rahonavis, ursprünglich als Frühaufsteher beschrieben,[70] wurde in mehreren Studien als nicht-aviäres Dromaeosaurid identifiziert.[63][71] Es wurde auch vorgeschlagen, dass Dromaeosauriden und Troodontiden selbst innerhalb von Aves und nicht nur außerhalb davon liegen.[72][73]
Eigenschaften, die Vögel und Dinosaurier verbinden[edit]
Viele anatomische[74] Merkmale werden von Vögeln und Theropodendinosauriern geteilt.
Gefieder[edit]
Archaeopteryx, das erste gute Beispiel eines “gefiederten Dinosauriers”, wurde 1861 entdeckt. Das erste Exemplar wurde im süddeutschen Solnhofener Kalkstein gefunden Lagerstätte, eine seltene und bemerkenswerte geologische Formation, die für ihre hervorragend detaillierten Fossilien bekannt ist. Archaeopteryx ist ein Übergangsfossil mit Merkmalen, die eindeutig zwischen denen von Nicht-Vogel-Theropod-Dinosauriern und Vögeln liegen. Entdeckt nur zwei Jahre nach Darwins wegweisendem Entstehung der ArtenDiese Entdeckung spornte die aufkommende Debatte zwischen Befürwortern der Evolutionsbiologie und des Kreationismus an. Dieser frühe Vogel ist so dinosaurierartig, dass ohne einen klaren Eindruck von Federn im umgebenden Felsen mindestens ein Exemplar verwechselt wurde Compsognathus.[75]


- Zahlen im Bild ausblenden
Schaufel
Nachfeder
Hohlwelle
(Calamus)
Seit den 1990er Jahren wurde eine Reihe weiterer gefiederter Dinosaurier gefunden, die die enge Beziehung zwischen Dinosauriern und modernen Vögeln noch deutlicher belegen. Die ersten davon wurden ursprünglich als einfach filamentös beschrieben Protofedern, die in Dinosaurierlinien berichtet wurden, die so primitiv sind wie Compsognathiden und Tyrannosauroiden.[76] Federn, die nicht von denen moderner Vögel zu unterscheiden waren, wurden jedoch bald darauf auch bei nicht-avialanischen Dinosauriern gefunden.[40]
Eine kleine Minderheit von Forschern hat behauptet, dass die einfachen filamentösen “Protofeder” -Strukturen einfach das Ergebnis der Zersetzung von Kollagenfasern unter der Haut der Dinosaurier oder in Flossen entlang ihres Rückens sind und dass es Arten mit unbestreitbaren Federn wie Oviraptorosauriern und Dromaeosauriern gibt keine Dinosaurier, sondern echte Vögel, die nichts mit Dinosauriern zu tun haben.[77] Eine Mehrheit der Studien hat jedoch festgestellt, dass gefiederte Dinosaurier tatsächlich Dinosaurier sind und dass die einfacheren Filamente unbestreitbarer Theropoden einfache Federn darstellen. Einige Forscher haben das Vorhandensein von farbtragendem Melanin in den Strukturen nachgewiesen – was bei Federn, aber nicht bei Kollagenfasern zu erwarten wäre.[78] Andere haben anhand von Studien zur modernen Vogelzersetzung gezeigt, dass selbst fortgeschrittene Federn unter den während der Fossilisierung auftretenden Quetschkräften filamentös erscheinen und dass die angeblichen “Protofedern” möglicherweise komplexer waren als bisher angenommen.[79] Detaillierte Untersuchung der “Protofedern” von Sinosauropteryx prima zeigten, dass einzelne Federn aus einer zentralen Feder bestanden (Rachis) mit Verdünner Widerhaken Abzweigung davon, ähnlich, aber primitiver in der Struktur als moderne Vogelfedern.[80]


Skelett[edit]
Da Federn oft mit Vögeln in Verbindung gebracht werden, werden gefiederte Dinosaurier oft als das fehlende Bindeglied zwischen Vögeln und Dinosauriern angepriesen. Die vielfältigen Skelettmerkmale, die auch die beiden Gruppen gemeinsam haben, stellen jedoch das wichtigere Bindeglied für Paläontologen dar. Darüber hinaus wird immer deutlicher, dass die Beziehung zwischen Vögeln und Dinosauriern und die Entwicklung des Fluges komplexere Themen sind als bisher angenommen. Während früher angenommen wurde, dass sich Vögel in einem linearen Verlauf aus Dinosauriern entwickelt haben, schließen einige Wissenschaftler, insbesondere Gregory S. Paul, dass Dinosaurier wie die Dromaeosaurier sich aus Vögeln entwickelt haben und die Flugkraft verlieren, während sie ihre Federn behalten in ähnlicher Weise wie der moderne Strauß und andere Laufvögel.
Vergleiche von Vogel- und Dinosaurierskeletten sowie kladistische Analysen bestätigen die Verbindung, insbesondere für einen Zweig von Theropoden, die Maniraptoren genannt werden. Skelettähnlichkeiten umfassen Hals, Schambein, Handgelenk (halbwahnsinniger Handwurzel), Arm und Brustgürtel, Schulterblatt, Schlüsselbein und Brustbein.
Eine Studie, in der embryonale, juvenile und adulte Archosaurierschädel verglichen wurden, kam zu dem Schluss, dass Vogelschädel durch Progenese, eine Art paedomorpher Heterochronie, von denen von Theropodendinosauriern abgeleitet wurden, was zur Beibehaltung der jugendlichen Eigenschaften ihrer Vorfahren führte.[81]
Lunge[edit]


Laut einer Untersuchung von Patrick M. O’Connor von der Ohio University hatten große fleischfressende Dinosaurier ein komplexes System von Luftsäcken, die denen moderner Vögel ähnelten. Bei Theropodendinosauriern (Fleischfressern, die auf zwei Beinen gingen und vogelähnliche Füße hatten) pumpten flexible Weichteil-Luftsäcke wahrscheinlich Luft durch die steifen Lungen, wie dies bei Vögeln der Fall ist. “Was früher formal als einzigartig für Vögel galt, war in irgendeiner Form bei den Vorfahren der Vögel vorhanden”, sagte O’Connor.[82][83]
Herz[edit]
Computertomographie (CT) -Scans im Jahr 2000 der Brusthöhle einer Probe des Ornithopoden Thescelosaurus fanden die offensichtlichen Überreste eines komplexen Herzens mit vier Kammern, ähnlich wie die heutigen Säugetiere und Vögel.[84] Die Idee ist innerhalb der wissenschaftlichen Gemeinschaft umstritten und wird als schlechte anatomische Wissenschaft kritisiert[85] oder einfach Wunschdenken.[86]
In einer 2011 veröffentlichten Studie wurden mehrere Untersuchungslinien auf die Frage nach der Identität des Objekts angewendet, darunter fortgeschrittenere CT-Scans, Histologie, Röntgenbeugung, Röntgenphotoelektronenspektroskopie und Rasterelektronenmikroskopie. Bei diesen Methoden stellten die Autoren fest, dass: die innere Struktur des Objekts keine Kammern enthält, sondern aus drei nicht verbundenen Bereichen aus Material geringerer Dichte besteht und nicht mit der Struktur des Herzens eines Straußes vergleichbar ist; Die “Wände” bestehen aus Sedimentmineralien, von denen nicht bekannt ist, dass sie in biologischen Systemen produziert werden, wie Goethit, Feldspatmineralien, Quarz und Gips sowie einigen Pflanzenfragmenten. Kohlenstoff, Stickstoff und Phosphor, chemische Elemente, die für das Leben wichtig sind, fehlten in ihren Proben; und Herzzellstrukturen fehlten. Es gab einen möglichen Fleck mit tierischen Zellstrukturen. Die Autoren fanden heraus, dass ihre Daten die Identifizierung als Konkretion von Sand aus der Grabumgebung und nicht aus dem Herzen unterstützten, mit der Möglichkeit, dass isolierte Gewebebereiche erhalten blieben.[87]
Die Frage, wie dieser Fund die Stoffwechselrate und die innere Anatomie des Dinosauriers widerspiegelt, ist jedoch ungeachtet der Identität des Objekts umstritten.[87] Sowohl moderne Krokodile als auch Vögel, die nächsten lebenden Verwandten von Dinosauriern, haben Vierkammerherzen (wenn auch in Krokodilen modifiziert), so dass Dinosaurier sie wahrscheinlich auch hatten; Die Struktur ist nicht unbedingt an die Stoffwechselrate gebunden.[88]
Schlafhaltung[edit]
Fossilien der Troodonten Mei und Sinornithoides zeigen, dass die Dinosaurier wie bestimmte moderne Vögel schliefen und ihre Köpfe unter den Armen steckten.[89] Dieses Verhalten, das möglicherweise dazu beigetragen hat, den Kopf warm zu halten, ist auch für moderne Vögel charakteristisch.
Reproduktionsbiologie[edit]
Beim Legen von Eiern wachsen weibliche Vögel eine besondere Art von Knochen in ihren Gliedern. Dieser Markknochen bildet eine kalziumreiche Schicht innerhalb des harten äußeren Knochens und wird als Kalziumquelle zur Herstellung von Eierschalen verwendet. Das Vorhandensein von endostal abgeleiteten Knochengeweben, die die inneren Markhöhlen von Teilen von a auskleiden Tyrannosaurus Rex Das hintere Glied des Exemplars deutete darauf hin T-Rex verwendeten ähnliche Fortpflanzungsstrategien und zeigten, dass die Probe weiblich ist.[90] Weitere Forschungen haben Markknochen im Theropoden gefunden Allosaurus und Ornithopod Tenontosaurus. Weil die Linie der Dinosaurier das beinhaltet Allosaurus und Tyrannosaurus weicht von der Linie ab, die zu führte Tenontosaurus Sehr früh in der Entwicklung der Dinosaurier deutet dies darauf hin, dass Dinosaurier im Allgemeinen Markgewebe produzieren.[91]
Brüten und Fürsorge für junge Menschen[edit]


Mehrere Citipati Es wurden Exemplare gefunden, die über den Eiern in ihrem Nest in einer Position ruhen, die am meisten an Brüten erinnert.[92]
Zum Beispiel zahlreiche Dinosaurierarten Maiasaurawurden in Herden gefunden, die sowohl sehr junge als auch erwachsene Individuen mischen, was auf reiche Wechselwirkungen zwischen ihnen hindeutet.
Es wurde ein Dinosaurierembryo ohne Zähne gefunden, was darauf hindeutet, dass eine gewisse elterliche Fürsorge erforderlich war, um den jungen Dinosaurier zu füttern. Möglicherweise spuckte der erwachsene Dinosaurier Nahrung in den Mund des jungen Dinosauriers aus (sehen altricial). Dieses Verhalten tritt bei zahlreichen Vogelarten auf; Elternvögel erbrechen das Futter im Maul des Jungtiers.
Muskelmagensteine[edit]
Sowohl Vögel als auch Dinosaurier verwenden Muskelmagensteine. Diese Steine werden von Tieren verschluckt, um die Verdauung zu unterstützen und Nahrung und harte Fasern abzubauen, sobald sie in den Magen gelangen. In Verbindung mit Fossilien werden Muskelmagen als Gastrolithen bezeichnet.[93]
Mägensteine kommen auch in einigen Fischen (Meeräsche, Schlammschatten und das Gillaroo, eine Art Forelle) und in Krokodilen vor.
Molekulare Beweise[edit]
Bei mehreren Gelegenheiten wurde die Extraktion von DNA und Proteinen aus Fossilien mesozoischer Dinosaurier behauptet, was einen Vergleich mit Vögeln ermöglicht. In Dinosaurierfossilien wurden vermutlich mehrere Proteine nachgewiesen.[94] einschließlich Hämoglobin.[95]
In der März 2005 Ausgabe von WissenschaftDr. Mary Higby Schweitzer und ihr Team kündigten die Entdeckung eines flexiblen Materials an, das dem tatsächlichen Weichgewebe eines 68-Millionen-Jährigen ähnelt Tyrannosaurus Rex Beinknochen der Probe MOR 1125 aus der Hell Creek Formation in Montana. Die sieben aus den Knochenfragmenten erhaltenen Kollagentypen im Vergleich zu Kollagendaten von lebenden Vögeln (insbesondere einem Huhn) legen nahe, dass ältere Theropoden und Vögel eng miteinander verwandt sind.[96] Das Weichgewebe ermöglichte einen molekularen Vergleich der Zellanatomie und der Proteinsequenzierung von Kollagengewebe, die 2007 veröffentlicht wurden. Beide zeigten dies an T-Rex und Vögel sind enger miteinander verwandt als beide Alligator.[97][98] Eine zweite molekulare Studie unterstützte die Beziehung von Vögeln zu Dinosauriern, obwohl sie erwartungsgemäß keine Vögel in Theropoda platzierte. Diese Studie verwendete acht zusätzliche Kollagensequenzen, die aus einem Femur des “mumifizierten” extrahiert wurden. Brachylophosaurus canadensis Probe MOR 2598, ein Hadrosaurier.[99] Diese Ergebnisse waren jedoch sehr kontrovers. Es wurden keine anderen Peptide im mesozoischen Alter berichtet. Im Jahr 2008 wurde vermutet, dass das vermutete Weichgewebe tatsächlich ein bakterieller Mikrofilm war.[100] Als Reaktion darauf wurde argumentiert, dass genau diese Mikrofilme das Weichgewebe schützten.[101] Ein weiterer Einwand war, dass die Ergebnisse durch Kontamination verursacht worden sein könnten.[102] Im Jahr 2015 wurden die Peptide unter kontrollierten Bedingungen, die vor Kontamination geschützt waren, noch identifiziert.[103] Im Jahr 2017 ergab eine Studie, dass im Knochen des modernen Straußes ein Peptid vorhanden war, das mit dem im Knochen identisch war Tyrannosaurus und Brachylophosaurus Proben, die die Gefahr einer Kreuzkontamination hervorheben.[104]
Über die erfolgreiche Extraktion alter DNA aus Dinosaurierfossilien wurde zweimal berichtet, aber bei weiterer Prüfung und Begutachtung durch Fachkollegen konnte keiner dieser Berichte bestätigt werden.[105]
Ursprung des Vogelfluges[edit]
Debatten über den Ursprung des Vogelflugs sind fast so alt wie die Idee, dass Vögel aus Dinosauriern entstanden sind, die kurz nach der Entdeckung von entstanden sind Archaeopteryx 1862. Zwei Theorien haben seitdem den größten Teil der Diskussion dominiert: Die Cursor-Theorie (“von Grund auf”) besagt, dass sich Vögel aus kleinen, schnellen Raubtieren entwickelt haben, die auf dem Boden liefen; Die Theorie der Baumbewohner (“von den Bäumen abwärts”) besagt, dass sich der Motorflug aus dem nicht angetriebenen Gleiten von Baumtieren (Baumklettern) entwickelt hat. Eine neuere Theorie, “Wing-Assisted Steigungslauf” (WAIR), ist eine Variante der Cursortheorie und schlägt vor, dass Flügel ihre aerodynamischen Funktionen aufgrund der Notwendigkeit entwickeln, sehr steile Hänge wie Bäume schnell hinaufzulaufen Hilf kleinen gefiederten Dinosauriern, vor Raubtieren zu fliehen.
Im März 2018 berichteten Wissenschaftler darüber Archaeopteryx war wahrscheinlich flugfähig, aber in einer Weise, die sich wesentlich von der moderner Vögel unterscheidet.[106][107]
Cursorial-Theorie (“von Grund auf”)[edit]


Die Cursortheorie des Ursprungs der Flucht wurde zuerst von Samuel Wendell Williston vorgeschlagen und von Baron Nopcsa ausgearbeitet. Diese Hypothese legt nahe, dass einige schnell laufende Tiere mit langen Schwänzen ihre Arme benutzten, um beim Laufen das Gleichgewicht zu halten. Moderne Versionen dieser Theorie unterscheiden sich in vielen Details von der Williston-Nopcsa-Version, hauptsächlich aufgrund von Entdeckungen seit Nopcsas Zeit.
Nopcsa vermutete, dass eine Vergrößerung der Oberfläche der ausgestreckten Arme kleinen Cursorial-Raubtieren helfen könnte, das Gleichgewicht zu halten, und dass sich die Schuppen der Unterarme verlängerten und sich zu Federn entwickelten. Die Federn könnten auch verwendet worden sein, um Insekten oder andere Beute zu fangen. Nach und nach sprangen die Tiere über größere Entfernungen, unterstützt von ihren sich entwickelnden Flügeln. Nopcsa schlug auch drei Stufen in der Entwicklung des Fluges vor. Zunächst entwickelten die Tiere einen passiven Flug, bei dem die Entwicklung von Flügelstrukturen als eine Art Fallschirm diente. Zweitens erreichten sie einen aktiven Flug, indem sie mit den Flügeln schlugen. Er verwendete Archaeopteryx als Beispiel für diese zweite Stufe. Schließlich erlangten die Vögel die Fähigkeit zu fliegen.[108]


Derzeit wird angenommen, dass sich Federn nicht aus Schuppen entwickelt haben, da Federn aus verschiedenen Proteinen bestehen.[109] Im Ernst, Nopcsas Theorie geht davon aus, dass sich Federn als Teil der Evolution des Fliegens entwickelt haben, und jüngste Entdeckungen beweisen, dass die Annahme falsch ist.
Federn sind bei Coelurosaurier-Dinosauriern (einschließlich des frühen Tyrannosauroids) sehr häufig Dilong).[110] Moderne Vögel werden von fast allen Paläontologen als Coelurosaurier eingestuft.[111] wenn auch nicht von einigen Ornithologen.[112][113][114] Die moderne Version der Hypothese “von Grund auf” argumentiert, dass die Vorfahren der Vögel klein waren, gefiedert, bodenlaufende räuberische Dinosaurier (eher wie Roadrunner in ihrem Jagdstil[115]) die ihre Vorderbeine für das Gleichgewicht verwendeten, während sie Beute verfolgten, und dass sich die Vorderbeine und Federn später auf eine Weise entwickelten, die Gleiten und dann Motorflug ermöglichte. Die am häufigsten vorgeschlagenen ursprünglichen Funktionen von Federn umfassen Wärmeisolierung und wettbewerbsfähige Anzeigen, wie bei modernen Vögeln.[116][117]
Alle der Archaeopteryx Fossilien stammen aus marinen Sedimenten, und es wurde vermutet, dass Flügel den Vögeln geholfen haben könnten, wie Wasser über Wasser zu laufen Jesus Christus Eidechse (Gemeiner Basilisk).[118]
Die jüngsten Widerlegungen der Hypothese “von Grund auf” versuchen, die Annahme der modernen Version zu widerlegen, dass Vögel modifizierte Coelurosaurier-Dinosaurier sind. Die stärksten Angriffe basieren auf embryologischen Analysen, aus denen hervorgeht, dass die Flügel der Vögel aus den Ziffern 2, 3 und 4 bestehen (entsprechend dem Zeige-, Mittel- und Ringfinger beim Menschen. Die erste der drei Ziffern eines Vogels bildet die Alula). die sie verwenden, um ein Abwürgen im Niedriggeschwindigkeitsflug zu vermeiden – zum Beispiel bei der Landung). Die Hände von Coelurosauriern werden jedoch durch die Ziffern 1, 2 und 3 (Daumen und die ersten beiden Finger beim Menschen) gebildet.[119] Diese embryologischen Analysen wurden jedoch sofort mit den embryologischen Gründen in Frage gestellt, dass sich die “Hand” in Kladen, die im Laufe ihrer Entwicklung einige Ziffern verloren haben, häufig anders entwickelt und dass sich die “Hände” der Vögel aus den Ziffern 1, 2 und 2 entwickeln 3.[120][121][122] Diese Debatte ist komplex und noch nicht gelöst – siehe “Digit Homology”.
Flügelunterstütztes Steigungslaufen[edit]
Die Hypothese des flügelunterstützten Steigungslaufs (WAIR) wurde durch die Beobachtung junger Chukar-Küken angeregt und schlägt vor, dass Flügel ihre aerodynamischen Funktionen entwickelten, weil sie sehr steile Hänge wie Baumstämme schnell hinauflaufen mussten, um beispielsweise zu entkommen Raubtiere.[123] Dies macht es zu einer speziellen Art von Cursor-Theorie (“von Grund auf”). Beachten Sie, dass in diesem Szenario Vögel benötigen Abtrieb um ihren Füßen mehr Halt zu geben.[124][125] Aber Frühaufsteher, einschließlich Archaeopteryxfehlte der Schultermechanismus, durch den moderne Vogelflügel schnelle, kraftvolle Aufschläge erzeugen. Da der Abtrieb, von dem WAIR abhängt, durch Aufschläge erzeugt wird, scheinen Frühaufsteher nicht in der Lage zu sein, WAIR auszuführen.[126] Da WAIR ein Verhaltensmerkmal ohne osteologische Spezialisierungen ist, ist es aufgrund der phylogenetischen Platzierung des Flugschlags vor der Divergenz der Neornithes, der Gruppe, die alle vorhandenen Vögel enthält, unmöglich festzustellen, ob WAIR Vorfahren des Vogelflugschlags ist oder von diesem abgeleitet ist .[127]
Baumtheorie (“von den Bäumen abwärts”)[edit]


Die meisten Versionen der Baumhypothese besagen, dass die Vorfahren der Vögel sehr kleine Dinosaurier waren, die in Bäumen lebten und von Ast zu Ast sprangen. Dieser kleine Dinosaurier hatte bereits Federn, die von der Evolution kooptiert wurden, um längere, steifere Formen zu erzeugen, die für die Aerodynamik nützlich waren und schließlich Flügel hervorbrachten. Wings hätten sich dann weiterentwickelt und als Vorrichtungen zunehmend verfeinert, um dem Springer mehr Kontrolle zu geben, Fallschirm zu springen, zu gleiten und schrittweise zu fliegen. Die Baumhypothese stellt auch fest, dass die Aerodynamik bei Baumtieren weitaus energieeffizienter ist, da solche Tiere einfach fallen, um minimale Gleitgeschwindigkeiten zu erreichen.[128][129]
Mehrere kleine Dinosaurier aus der Jura- oder frühen Kreidezeit, alle mit Federn, wurden so interpretiert, dass sie möglicherweise baumartige und / oder aerodynamische Anpassungen aufweisen. Diese schließen ein Scansoriopteryx, Epidexipteryx, Microraptor, Pedopenna, und Anchiornis. Anchiornis ist für dieses Thema besonders wichtig, da es lange zuvor zu Beginn des späten Jura lebte Archaeopteryx.[130]
Analyse der Proportionen der Zehenknochen der primitivsten Vögel Archaeopteryx und Konfuziusornisdeuten im Vergleich zu lebenden Arten darauf hin, dass die frühen Arten möglicherweise sowohl am Boden als auch in Bäumen gelebt haben.[131]
Eine Studie ergab, dass die frühesten Vögel und ihre unmittelbaren Vorfahren keine Bäume bestiegen. Diese Studie ergab, dass das Ausmaß der Krümmung der Zehenklauen bei Frühaufstehern eher dem bei modernen Bodensuchvögeln als bei Sitzvögeln entspricht.[132]
Verminderte Bedeutung von Archaeopteryx[edit]
Archaeopteryx war das erste und lange Zeit das einzige bekannte gefiederte mesozoische Tier. Infolgedessen konzentrierte sich die Diskussion über die Entwicklung der Vögel und des Vogelfluges auf Archaeopteryx zumindest bis Mitte der neunziger Jahre.

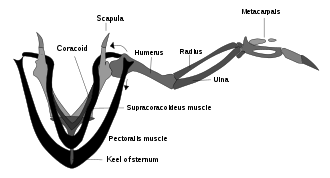
Es gab Debatten darüber, ob Archaeopteryx könnte wirklich fliegen. Anscheinend Archaeopteryx hatte die Gehirnstrukturen und Innenohr-Gleichgewichtssensoren, mit denen Vögel ihren Flug steuern.[133]Archaeopteryx hatte auch eine Flügelfederanordnung wie die moderner Vögel und ähnlich asymmetrische Flugfedern an Flügeln und Schwanz. Aber Archaeopteryx fehlte der Schultermechanismus, durch den die Flügel moderner Vögel schnelle, kraftvolle Aufschläge erzeugen (siehe Abbildung oben von der Supracoracoideus-Riemenscheibe); Dies könnte bedeuten, dass es und andere Frühaufsteher nicht in der Lage waren zu fliegen und nur gleiten konnten.[126]
Aber die Entdeckung vieler gefiederter Dinosaurier seit Anfang der neunziger Jahre bedeutet dies Archaeopteryx ist nicht länger die Schlüsselfigur in der Entwicklung des Vogelfluges. Andere kleine gefiederte Coelurosaurier aus der Kreidezeit und dem späten Jura zeigen mögliche Vorläufer des Vogelfluges. Diese schließen ein Rahonavis, ein Groundrunner mit a Velociraptor– wie eine erhabene Sichelklaue am zweiten Zeh, von der einige Paläontologen annehmen, dass sie besser für den Flug geeignet ist als Archaeopteryx,[134]Scansoriopteryx, ein Baumdinosaurier, der die Theorie “von den Bäumen abwärts” unterstützen könnte,[135] und MicroraptorEin Baumdinosaurier, der möglicherweise zum Motorflug fähig ist, aber in diesem Fall eher einem Doppeldecker ähnelt, da er gut entwickelte Federn an den Beinen hat.[136] Bereits 1915 argumentierten einige Wissenschaftler, dass die Entwicklung des Vogelfluges einen vierflügeligen (oder Tetrapteryx) Bühne.[137][138]
Sekundäre Flugunfähigkeit bei Dinosauriern[edit]
Gruppen, die normalerweise als Vögel angesehen werden, sind fett gedruckt.[73]
Eine Hypothese, die Gregory Paul gutgeschrieben und in seinen Büchern vorgeschlagen wurde Raubdinosaurier der Welt (1988) und Dinosaurier der Luft (2002) legen nahe, dass einige Gruppen nicht fliegender fleischfressender Dinosaurier – insbesondere Deinonychosaurier, aber möglicherweise auch andere wie Oviraptorosaurier, Therizinosaurier, Alvarezsauriden und Ornithomimosaurier – tatsächlich von Vögeln abstammen. Paul schlug auch vor, dass der Vogelvorfahr dieser Gruppen in seinen Fluganpassungen weiter fortgeschritten war als Archaeopteryx. Die Hypothese würde das bedeuten Archaeopteryx ist weniger eng mit vorhandenen Vögeln verwandt als diese Dinosaurier.[139]
Die Hypothese von Paulus erhielt zusätzliche Unterstützung, als Mayr et al. (2005) analysierten ein neues, zehntes Exemplar von Archaeopteryxund kam zu dem Schluss Archaeopteryx war die Schwesterklade der Deinonychosauria, aber das war der fortgeschrittenere Vogel Konfuziusornis war innerhalb der Dromaeosauridae. Dieses Ergebnis stützt die Hypothese von Paulus, wonach die Deinonychosauria und die Troodontidae Teil von Aves sind, der eigentlichen Vogellinie, und zweitens flugunfähig. Dieses Papier schloss jedoch alle anderen Vögel aus und untersuchte daher nicht ihre Charakterverteilungen. Das Papier wurde von Corfe und Butler (2006) kritisiert, die feststellten, dass die Autoren ihre Schlussfolgerungen statistisch nicht stützen konnten. Mayr et al. stimmte zu, dass die statistische Unterstützung schwach war, fügte jedoch hinzu, dass sie auch für die alternativen Szenarien schwach ist.[141]
Aktuelle kladistische Analysen stützen Pauls Hypothese über die Position von nicht Archaeopteryx. Stattdessen zeigen sie das an Archaeopteryx ist näher an Vögeln, innerhalb der Klade Avialae, als es zu Deinonychosauriern oder Oviraptorosauriern ist. Einige Fossilien unterstützen jedoch die Version dieser Theorie, wonach einige nicht fliegende fleischfressende Dinosaurier möglicherweise fliegende Vorfahren hatten. Speziell, Microraptor, Pedopenna, und Anchiornis Alle haben geflügelte Füße, teilen viele Merkmale und liegen nahe der Basis der Clade Paraves. Dies deutet darauf hin, dass der Ahnenparavian ein vierflügeliger Segelflugzeug war und dass größere Deinonychosaurier sekundär die Gleitfähigkeit verloren, während die aerodynamische Fähigkeit der Vogellinie im Laufe der Zeit zunahm.[2]Deinonychus kann auch eine teilweise Volanz aufweisen, wobei die Jungen flug- oder gleitfähig sind und die Erwachsenen flugunfähig sind.[142] Im Jahr 2018 kam eine Studie zu dem Schluss, dass der letzte gemeinsame Vorfahr der Pennaraptora Gelenkflächen an den Fingern sowie zwischen Mittelfuß und Handgelenk hatte, die zur Stabilisierung der Hand im Flug optimiert wurden. Dies wurde als Hinweis auf sekundäre Flugunfähigkeit bei schweren Basalmitgliedern dieser Gruppe angesehen.[143]
In Euornithes ist das früheste eindeutige Beispiel für sekundäre Flugunfähigkeit Patagopteryx.[144]
Siehe auch[edit]
![]()

- ^ Chiappe, Luis M. (2009). “Verkleinerte Dinosaurier: Der evolutionäre Übergang zu modernen Vögeln”. Evolution: Bildung und Öffentlichkeitsarbeit. 2 (2): 248–256. doi:10.1007 / s12052-009-0133-4.
- ^ ein b Witmer, LM (2009). “Paläontologie: Gefiederte Dinosaurier in einem Gewirr”. Natur. 461 (7264): 601–2. Bibcode:2009Natur.461..601W. doi:10.1038 / 461601a. PMID 19794481. S2CID 205049989.
- ^ Darwin, Charles R. (1859). Über die Entstehung von Arten durch natürliche Selektion oder die Erhaltung bevorzugter Rassen im Kampf ums Leben. London: John Murray. p. 502pp. ISBN 978-1-4353-9386-8.
- ^ von Meyer, CE Hermann. (1861). “”Archaeopteryx lithographica (Vogel-Feder) und Pterodactylus von Solnhofen “. Neues Jahrbuch für Mineralogie, Geologie und Paläontologie (auf Deutsch). 1861: 678–679.
- ^ Owen, Richard. (1863). “Auf der Archaeopteryx [sp] von Meyer mit einer Beschreibung der fossilen Überreste einer Langschwanzart aus dem lithografischen Stein von Solenhofen [sp]””. Philosophische Transaktionen der Royal Society of London. 153: 33–47. doi:10.1098 / rstl.1863.0003.
- ^ Gegenbaur, K. (1863). “Vergleichend-anatomische Bemerkungen über das Fußskelet der Vögel”. Archiv für Anatomie, Physiologie und Wissenschaftliche Medizin. 1863: 450–472.
- ^ Cope, ED (1867). “Ein Bericht über die ausgestorbenen Reptilien, die sich den Vögeln näherten”. Verfahren der Akademie der Naturwissenschaften von Philadelphia. 19: 234–235.Verknüpfung
- ^ Huxley, TH (1868). “Auf den Tieren, die fast zwischen Vögeln und Reptilien liegen”. Annalen und Zeitschrift für Naturgeschichte. 4 .. 2: 66–75.
- ^ Huxley, Thomas H. (1870). “Ein weiterer Beweis für die Affinität zwischen den Dinosaurier-Reptilien und den Vögeln”. Vierteljährliches Journal der Geological Society of London. 26 (1–2): 12–31. doi:10.1144 / GSL.JGS.1870.026.01-02.08. S2CID 129125487.
- ^ Nopcsa, Franz. (1907). “Ideen zum Ursprung des Fluges”. Verfahren der Zoological Society of London: 223–238.
- ^ Seeley, Harry G. (1901). Drachen der Luft: Ein Bericht über ausgestorbene fliegende Reptilien. London: Methuen & Co. p. 239pp.
- ^ Abel, O (1911). “Die gleichen der Vögel und ihre Lebensweise”. Verhandlungen der Zoologisch-Botanischen Gesellschaft in Wien. 61: 144–191.
- ^ Nieuwland, Ilja JJ (2004). “Gerhard Heilmann und das Auge des Künstlers in der Wissenschaft, 1912-1927” (PDF). PalArch’s Journal of Vertebrate Palaeontology. 3 (2). Archiviert von das Original (PDF) am 24.02.2016. Abgerufen 2009-05-21.
- ^ Heilmann, Gerhard (1926). Der Ursprung der Vögel. London: Witherby. p. 208pp. ISBN 978-0-486-22784-9.
- ^ ein b c Padian, Kevin. (2004). “Basal Avialae”. In Weishampel, David B.; Dodson, Peter; Osmólska, Halszka (Hrsg.). Die Dinosaurier (Zweite Ausgabe). Berkeley: University of California Press. S. 210–231. ISBN 978-0-520-24209-8.
- ^ Zum Beispiel fand Roy Chapman Andrews 1923, drei Jahre vor Heilmans ‘Buch, eine gute Oviraptor fossil in MongoleiHenry Fairfield Osborn, der das Fossil 1924 analysierte, identifizierte die Furcula fälschlicherweise als Interklavikel. beschrieben in Paul, GS (2002). Dinosaurier der Luft: Die Entwicklung und der Verlust des Fluges bei Dinosauriern und Vögeln. JHU Drücken Sie. ISBN 978-0-8018-6763-7.
- ^ Camp, Charles L. (1936). “Eine neue Art von kleinen Theropodendinosauriern aus dem Navajo-Sandstein von Arizona”. Bulletin des Instituts für Geologische Wissenschaften der Universität von Kalifornien. 24: 39–65.
- ^ In einem (n Oviraptor:: Barsbold, R. (1983). “Fleischfressende Dinosaurier aus der Kreidezeit der Mongolei”. Trudy Soumestnaya Sovetsko-Mongol’skaya Paleontogicheskaya Ekspeditsiya (auf Russisch). 19: 1–117. Siehe die Zusammenfassung und Bilder unter “Ein Wunsch für Coelophysis“”. Archiviert von das Original am 3. Februar 2008.
- ^ Lipkin, C., Sereno, PC und Horner, JR (November 2007). “Die Furcula in Suchomimus Tenerensis und Tyrannosaurus Rex (Dinosauria: Theropoda: Tetanurae)”. Zeitschrift für Paläontologie. 81 (6): 1523–1527. doi:10.1666 / 06-024.1. S2CID 86234363.CS1-Wartung: mehrere Namen: Autorenliste (Link) – Volltext derzeit online unter “Die Furcula in Suchomimus Tenerensis und Tyrannosaurus Rex“”. 2007-11-11. Archiviert von das Original am 22.05.2011. Abgerufen 2008-04-17. Dies listet eine große Anzahl von Theropoden auf, in denen Furculae gefunden wurden, und beschreibt diejenigen von Suchomimus Tenerensis und Tyrannosaurus Rex.
- ^ Carrano, MR, Hutchinson, JR und Sampson, SD (Dezember 2005). “Neue Informationen zu Segisaurus halli, ein kleiner Theropodendinosaurier aus dem frühen Jura von Arizona “ (PDF). Journal of Vertebrate Paleontology. 25 (4): 835–849. doi:10.1671 / 0272-4634 (2005) 025[0835:NIOSHA]2.0.CO; 2. ISSN 0272-4634. Archiviert von das Original (PDF) am 12.05.2006.CS1-Wartung: mehrere Namen: Autorenliste (Link)
- ^ Yates, Adam M.; und Vasconcelos, Cecilio C. (2005). “Furcula-ähnliche Schlüsselbeine im Prosauropodendinosaurier Massospondylus“. Journal of Vertebrate Paleontology. 25 (2): 466–468. doi:10.1671 / 0272-4634 (2005) 025[0466:FCITPD]2.0.CO; 2. ISSN 0272-4634.
- ^ Ostrom, John H. (1969). “Osteologie von Deinonychus antirrhopus, ein ungewöhnlicher Theropod aus der Unterkreide von Montana “. Bulletin des Peabody Museum of Natural History. 30: 1–165.
- ^ Ostrom, John H. (1970). “”Archaeopteryx: Hinweis auf ein “neues” Exemplar “. Wissenschaft. 170 (3957): 537–8. Bibcode:1970Sci … 170..537O. doi:10.1126 / science.170.3957.537. PMID 17799709. S2CID 46682559.
- ^ Chambers, Paul (2002). Streitpunkte: Die Archaeopteryx-Skandale. London: John Murray Ltd., S. 183–184. ISBN 978-0-7195-6054-5.
- ^ Walker, Alick D. (1972). “Neues Licht auf die Herkunft von Vögeln und Krokodilen”. Natur. 237 (5353): 257–263. Bibcode:1972Natur.237..257W. doi:10.1038 / 237257a0. S2CID 4174690.
- ^ Ostrom, John H. (1973). “Die Abstammung der Vögel”. Natur. 242 (5393): 136. Bibcode:1973NPhS..242..136O. doi:10.1038 / 242136a0. S2CID 29873831.
- ^ Ostrom, John H. (1975). Der Ursprung der Vögel. Jahresrückblick auf die Erd- und Planetenwissenschaften. 3. S. 55–77. Bibcode:1975AREPS … 3 … 55O. doi:10.1146 / annurev.ea.03.050175.000415. ISBN 978-0-912532-57-8.
- ^ Ostrom, John H. (1976). “”Archaeopteryx und der Ursprung der Vögel “. Biologisches Journal der Linnean Society. 8 (2): 91–182. doi:10.1111 / j.1095-8312.1976.tb00244.x.
- ^ Bakker, Robert T. (1972). “Anatomische und ökologische Hinweise auf Endothermie bei Dinosauriern”. Natur. 238 (5359): 81–85. Bibcode:1972Natur.238 … 81B. doi:10.1038 / 238081a0. S2CID 4176132.
- ^ Horner, John R.; Makela, Robert (1979). “Das Nest der Jugendlichen liefert Hinweise auf die Familienstruktur der Dinosaurier”. Natur. 282 (5736): 296–298. Bibcode:1979Natur.282..296H. doi:10.1038 / 282296a0. S2CID 4370793.
- ^ Hennig, EH Willi (1966). Phylogenetische Systematik. übersetzt von Davis, D. Dwight; & Zangerl, Rainer. Urbana: University of Illinois Press. ISBN 978-0-252-06814-0.
- ^ ein b Gauthier, Jacques. (1986). “Saurischian Monophyly und der Ursprung der Vögel”. In Padian Kevin (Hrsg.). Der Ursprung der Vögel und die Entwicklung des Fluges. Memoiren der kalifornischen Akademie der Wissenschaften 8. S. 1–55.
- ^ ein b c d Senter, Phil (2007). “Ein neuer Blick auf die Phylogenie von Coelurosauria (Dinosauria: Theropoda)”. Zeitschrift für systematische Paläontologie. 5 (4): 429–463. doi:10.1017 / S1477201907002143. S2CID 83726237.
- ^ ein b c Turner, Alan H.; Hwang, sonnig; & Norell, Mark A. (2007). “Ein kleiner Theropod aus Öösh, frühe Kreidezeit, Baykhangor, Mongolei”. American Museum Novitates. 3557 (1): 1–27. doi:10.1206 / 0003-0082 (2007) 3557[1:ASDTFS]2.0.CO; 2. hdl:2246/5845. ISSN 0003-0082.CS1-Wartung: mehrere Namen: Autorenliste (Link)
- ^ Sereno, Paul C.; Rao Chenggang (1992). “Frühe Entwicklung der Vogelflucht und des Sitzens: neue Erkenntnisse aus der Unterkreide Chinas”. Wissenschaft. 255 (5046): 845–8. Bibcode:1992Sci … 255..845S. doi:10.1126 / science.255.5046.845. PMID 17756432. S2CID 41106677.
- ^ Hou Lian-Hai, Lian-hai; Zhou Zhonghe; Martin, Larry D.; & Feduccia, Alan (1995). “Ein Schnabelvogel aus dem Jura Chinas”. Natur. 377 (6550): 616–618. Bibcode:1995Natur.377..616H. doi:10.1038 / 377616a0. S2CID 4357707.CS1-Wartung: mehrere Namen: Autorenliste (Link)
- ^ Ji Qiang; Ji Shu-an (1996). “Über die Entdeckung des frühesten Vogelfossils in China und die Herkunft der Vögel” (PDF). Chinesische Geologie. 233: 30–33.
- ^ Chen Pei-ji, Pei-ji; Dong Zhiming; & Zhen Shuo-nan. (1998). “Ein außergewöhnlich erhaltener Theropodendinosaurier aus der Yixian Formation of China”. Natur. 391 (6663): 147–152. Bibcode:1998Natur.391..147C. doi:10.1038 / 34356. S2CID 4430927.CS1-Wartung: mehrere Namen: Autorenliste (Link)
- ^ Lingham-Soliar, Theagarten; Feduccia, Alan; & Wang Xiaolin. (2007). “Ein neues chinesisches Exemplar weist darauf hin, dass ‘Protofedern’ im frühen Kreide-Theropodendinosaurier Sinosauropteryx abgebaute Kollagenfasern sind.”. Verfahren der Royal Society B.. 274 (1620): 1823–9. doi:10.1098 / rspb.2007.0352. PMC 2270928. PMID 17521978.CS1-Wartung: mehrere Namen: Autorenliste (Link)
- ^ ein b Ji Qiang, Philip J.; Currie, Philip J.; Norell, Mark A.; & Ji Shu-an. (1998). “Zwei gefiederte Dinosaurier aus Nordostchina” (PDF). Natur. 393 (6687): 753–761. Bibcode:1998Natur.393..753Q. doi:10.1038 / 31635. S2CID 205001388.CS1-Wartung: mehrere Namen: Autorenliste (Link)
- ^ Sloan, Christopher P. (1999). “Federn für T-Rex? “. National Geographic. 196 (5): 98–107. Archiviert von das Original am 11.10.2012.
- ^ Monastersky, Richard (2000). “Alles durcheinander mit Vögeln und Dinosauriern”. Wissenschaftsnachrichten. 157 (3): 38. doi:10.2307 / 4012298. JSTOR 4012298. Archiviert von das Original am 03.10.2008. Abgerufen 2008-07-04.
- ^ Xu Xing, Xing; Tang Zhi-lu; & Wang Xiaolin. (1999). “Ein therizinosaurider Dinosaurier mit integumentären Strukturen aus China”. Natur. 399 (6734): 350–354. Bibcode:1999Natur.399..350X. doi:10.1038 / 20670. S2CID 204993327.CS1-Wartung: mehrere Namen: Autorenliste (Link)
- ^ Xu Xing, X; Norell, Mark A.; Kuang Xuewen; Wang Xiaolin; Zhao Qi; & Jia Chengkai. (2004). “Basale Tyrannosauroide aus China und Hinweise auf Protofedern bei Tyrannosauroiden”. Natur. 431 (7009): 680–4. Bibcode:2004Natur.431..680X. doi:10.1038 / nature02855. PMID 15470426. S2CID 4381777.CS1-Wartung: mehrere Namen: Autorenliste (Link)
- ^ Xu Xing, X; Zhou Zhonghe; Wang Xiaolin; Kuang Xuewen; Zhang Fucheng; & Du Xiangke (2003). “Vierflügelige Dinosaurier aus China”. Natur. 421 (6921): 335–40. Bibcode:2003Natur.421..335X. doi:10.1038 / nature01342. PMID 12540892. S2CID 1160118.CS1-Wartung: mehrere Namen: Autorenliste (Link)
- ^ Zhou, Zhonghe; Zhang Fucheng (2002). “Ein langschwänziger, samenfressender Vogel aus der frühen Kreidezeit Chinas”. Natur. 418 (6896): 405–9. Bibcode:2002Natur.418..405Z. doi:10.1038 / nature00930. PMID 12140555. S2CID 4423299.
- ^ ein b Chatterjee, Sankar (17. April 1998). “Die Finger von Vögeln und Dinosauriern zählen”. Wissenschaft. 280 (5362): 355a – 355. Bibcode:1998Sci … 280..355C. doi:10.1126 / science.280.5362.355a.
- ^ Homepage der Abteilung für Geologie der Universität von Maryland, “Theropoda I” auf Avetheropoda, 14. Juli 2006.
- ^ Wagner, GP; Gautthier, JA (1999). “1,2,3 = 2,3,4: Eine Lösung für das Problem der Homologie der Ziffern in der Vogelhand”. Proc. Natl. Acad. Sci. Vereinigte Staaten von Amerika. 96 (9): 5111–6. Bibcode:1999PNAS … 96.5111W. doi:10.1073 / pnas.96.9.5111. PMC 21825. PMID 10220427.
- ^ Scienceblogs: Limusaurus ist großartig Archiviert 22.06.2009 an der Wayback-Maschine.
- ^ Entwicklungsbiologie 8e Online. Kapitel 16: Haben sich Vögel aus den Dinosauriern entwickelt? Archiviert 2008-01-18 an der Wayback-Maschine
- ^ Vargas AO, Wagner GP und Gauthier, JA. 2009. Identität von Limusaurus und Vogelziffern. Erhältlich bei Nature Precedings [1]
- ^ Havstad, Joyce C.; Smith, N. Adam (2019). “Fossilien mit Federn und Wissenschaftstheorie”. Systematische Biologie. 68 (5): 840–851. doi:10.1093 / sysbio / syz010. ISSN 1076-836X. PMC 6701454. PMID 30753719.
- ^ Martin, Larry D. (2004). “Ein basaler archosaurischer Ursprung für Vögel”. Acta Zoologica Sinica. 50 (6): 978–990.
- ^ Feduccia, Alan; Lingham-Soliar, Theagarten; & Hincliffe, J. Richard. (2005). “Gibt es gefiederte Dinosaurier? Testen der Hypothese auf neontologische und paläontologische Beweise”. Zeitschrift für Morphologie. 266 (2): 125–66. doi:10.1002 / jmor.10382. PMID 16217748. S2CID 15079072.CS1-Wartung: mehrere Namen: Autorenliste (Link)
- ^ Burke, Ann C.; Feduccia, Alan (1997). “Entwicklungsmuster und die Identifizierung von Homologien in der Vogelhand”. Wissenschaft. 278 (5338): 666–668. Bibcode:1997Sci … 278..666B. doi:10.1126 / science.278.5338.666.
- ^ Newman SA (2011). “Thermogenese, Muskelhyperplasie und der Ursprung der Vögel”. BioEssays. 33 (9): 653–656. doi:10.1002 / bies.201100061. PMID 21695679. S2CID 42012034.
- ^ Newman SA, Mezentseva NV, Badyaev AV (2013). “Genverlust, Thermogenese und Herkunft der Vögel”. Annalen der New Yorker Akademie der Wissenschaften. 1289 (1): 36–47. Bibcode:2013NYASA1289 … 36N. doi:10.1111 / nyas.12090. PMID 23550607. S2CID 12240405.
- ^ Mezentseva NV, Kumaratilake JS, Newman SA (2008). “Der Differenzierungsweg der braunen Adipozyten bei Vögeln: Ein Evolutionsweg, der nicht eingeschlagen wurde”. BMC Biology. 6 (1): 17. doi:10.1186 / 1741-7007-6-17. PMC 2375860. PMID 18426587.
- ^ Sereno, Paul C. (1997). “Der Ursprung und die Entwicklung der Dinosaurier”. Jahresrückblick auf die Erd- und Planetenwissenschaften. 25 (1): 435–489. Bibcode:1997AREPS..25..435S. doi:10.1146 / annurev.earth.25.1.435.
- ^ Chiappe, Luis M. (1997). “Aves”. In Currie, Philip J.; Padian, Kevin (Hrsg.). Enzyklopädie der Dinosaurier. San Diego: Akademische Presse. S. 45–50. ISBN 978-0-12-226810-6.
- ^ ein b Holtz, Thomas R.; Osmólska, Halszka. (2004). “Saurischia”. In Weishampel, David B.; Dodson, Peter; Osmólska, Halszka (Hrsg.). Die Dinosaurier (Zweite Ausgabe). Berkeley: University of California Press. S. 21–24. ISBN 978-0-520-24209-8.
- ^ ein b Turner, Alan H.; Pol, Diego; Clarke, Julia A.; Erickson, Gregory M.; & Norell, Mark A. (2007). “Eine basale Dromaeosaurier- und Größenentwicklung vor dem Vogelflug”. Wissenschaft. 317 (5843): 1378–81. Bibcode:2007Sci … 317.1378T. doi:10.1126 / science.1144066. PMID 17823350.CS1-Wartung: mehrere Namen: Autorenliste (Link)
- ^ Osmólska, Halszka; Maryańska, Teresa; & Wolsan, Mieczysław. (2002). “Avialan Status für Oviraptorosauria”. Acta Palaeontologica Polonica. 47 (1): 97–116.CS1-Wartung: mehrere Namen: Autorenliste (Link)
- ^ Martinelli, Agustín G.; Vera, Ezequiel I. (2007). “”Achillesaurus manazzonei, ein neuer alvarezsauridischer Theropod (Dinosauria) aus der spätkreidezeitlichen Bajo de la Carpa-Formation, Provinz Río Negro, Argentinien “ (PDF). Zootaxa. 1582: 1–17. doi:10.11646 / zootaxa.1582.1.1.
- ^ Novas, Fernando E.; Pol, Diego. (2002). “Alvarezsaurid Beziehungen überdacht”. In Chiappe, Luis M.; Witmer, Lawrence M. (Hrsg.). Mesozoische Vögel: Über den Köpfen der Dinosaurier. Berkeley: University of California Press. S. 121–125. ISBN 978-0-520-20094-4.
- ^ Sereno, Paul C. (1999). “Die Entwicklung der Dinosaurier”. Wissenschaft. 284 (5423): 2137–47. doi:10.1126 / science.284.5423.2137. PMID 10381873.
- ^ Perle, Altangerel; Norell, Mark A.; Chiappe, Luis M.; & Clark, James M. (1993). “Flugunfähiger Vogel aus der Kreidezeit der Mongolei”. Natur. 362 (6421): 623–626. Bibcode:1993Natur.362..623A. doi:10.1038 / 362623a0. S2CID 4252852.CS1-Wartung: mehrere Namen: Autorenliste (Link)
- ^ Chiappe, Luis M.; Norell, Mark A.; & Clark, James M. (2002). “Die kreidezeitlichen, kurzarmigen Alvarezsauridae: Mononykus und seine Verwandten “. In Chiappe, Luis M.; Witmer, Lawrence M. (Hrsg.). Mesozoische Vögel: Über den Köpfen der Dinosaurier. Berkeley: University of California Press. S. 87–119. ISBN 978-0-520-20094-4.CS1-Wartung: mehrere Namen: Autorenliste (Link)
- ^ Forster, Catherine A.; Sampson, Scott D.; Chiappe, Luis M.; & Krause, David W. (1998). “Die Theropoden-Abstammung der Vögel: neue Beweise aus der späten Kreidezeit Madagaskars”. Wissenschaft. 279 (5358): 1915–9. Bibcode:1998Sci … 279.1915F. doi:10.1126 / science.279.5358.1915. PMID 9506938.CS1-Wartung: mehrere Namen: Autorenliste (Link)
- ^ Makovicky, Peter J.; Apesteguía, Sebastián; & Agnolín, Federico L. (2005). “Der früheste dromaeosauride Theropod aus Südamerika”. Natur. 437 (7061): 1007–11. Bibcode:2005Natur.437.1007M. doi:10.1038 / nature03996. PMID 16222297. S2CID 27078534.CS1-Wartung: mehrere Namen: Autorenliste (Link)
- ^ Paul, Gregory S. (2002). Dinosaurier der Luft: Die Entwicklung und der Verlust des Fluges bei Dinosauriern und Vögeln. Baltimore: Johns Hopkins University Press. p. 472pp. ISBN 978-0-8018-6763-7.
- ^ ein b Mayr, Gerald; Pohl, Burkhard; & Peters, D. Stefan (2005). “Eine gut erhaltene Archaeopteryx Probe mit Theropodenmerkmalen “. Wissenschaft. 310 (5753): 1483–6. Bibcode:2005Sci … 310.1483M. doi:10.1126 / science.1120331. PMID 16322455. S2CID 28611454.CS1-Wartung: mehrere Namen: Autorenliste (Link)
- ^ Chatterjee, Immoor; L. Immoor (9. September 2005). “Die Dinosaurier der Jurassic Park Filme”. Geolor.com. Archiviert von das Original am 12. Mai 2008. Abgerufen 23. Juni 2007.
- ^ Wellnhofer, P. (1988). “Ein neuer Exemplar von Archaeopteryx“. Archaeopteryx. 6: 1–30.
- ^ Xu X, Norell MA, Kuang X, Wang X, Zhao Q, Jia C (Oktober 2004). “Basale Tyrannosauroide aus China und Hinweise auf Protofedern bei Tyrannosauroiden”. Natur. 431 (7009): 680–4. Bibcode:2004Natur.431..680X. doi:10.1038 / nature02855. PMID 15470426. S2CID 4381777.
- ^ Feduccia, A. (2012). Rätsel der gefiederten Drachen: Versteckte Vögel Chinas. Yale University Press, ISBN 0-300-16435-1, ISBN 978-0-300-16435-0
- ^ Zhang, F.; Kearns, SL; Orr, PJ; Benton, MJ; Zhou, Z.; Johnson, D.; Xu, X.; und Wang, X. (2010). “Versteinerte Melanosomen und die Farbe von Kreide-Dinosauriern und Vögeln” (PDF). Natur. 463 (7284): 1075–1078. Bibcode:2010Natur.463.1075Z. doi:10.1038 / nature08740. PMID 20107440. S2CID 205219587.CS1-Wartung: mehrere Namen: Autorenliste (Link)
- ^ Foth, C (2012). “Zur Identifizierung von Federstrukturen in Stammlinienvertretern von Vögeln: Beweise aus Fossilien und Aktuopalaeontologie”. Paläontologische Zeitschrift. 86: 91–102. doi:10.1007 / s12542-011-0111-3. S2CID 86362907.
- ^ Currie, PJ; Chen, P.-j. (2001). “Anatomie von Sinosauropteryx prima aus Liaoning, Nordostchina “. Kanadisches Journal of Earth Sciences. 38 (1): 705–727. Bibcode:2001CaJES..38.1705C. doi:10.1139 / cjes-38-12-1705.
- ^ Bhullar, BAS; Marugán-Lobón, JS; Racimo, F.; Bever, GS; Rowe, TB; Norell, MA; Abzhanov, A. (2012). “Vögel haben paedomorphe Dinosaurierschädel”. Natur. 487 (7406): 223–226. Bibcode:2012Natur.487..223B. doi:10.1038 / nature11146. PMID 22722850. S2CID 4370675.
- ^ O’Connor, PM; Claessens, LPAM (2005). “Grundlegendes Design der Vogellungen und Durchflussbeatmung bei Nicht-Vogel-Theropodendinosauriern”. Natur. 436 (7048): 253–6. Bibcode:2005Natur.436..253O. doi:10.1038 / nature03716. PMID 16015329. S2CID 4390587.
- ^ CS1-Wartung: mehrere Namen: Autorenliste (Link)
- ^ Fisher, PE; Russell, DA; Stoskopf, MK; Barrick, RE; Hammer, M.; Kuzmitz, AA (2000). “Kardiovaskuläre Hinweise auf eine mittlere oder höhere Stoffwechselrate bei einem ornithischianischen Dinosaurier”. Wissenschaft. 288 (5465): 503–5. Bibcode:2000Sci … 288..503F. doi:10.1126 / science.288.5465.503. PMID 10775107.
- ^ Hillenius, WJ; Ruben, JA (2004). “Die Entwicklung der Endothermie bei Landwirbeltieren: Wer? Wann? Warum?” Physiologische und biochemische Zoologie. 77 (6): 1019–42. doi:10.1086 / 425185. PMID 15674773. S2CID 29300018.
- ^ Rowe, T.; McBride, EF; Sereno, PC; Russell, DA; Fisher, PE; Barrick, RE; Stoskopf, MK (2001). “Dinosaurier mit Herz aus Stein”. Wissenschaft. 291 (5505): 783. doi:10.1126 / science.291.5505.783a. PMID 11157158.
- ^ ein b Cleland, Timothy P.; Stoskopf, Michael K.; und Schweitzer, Mary H. (2011). “Histologische, chemische und morphologische Nachuntersuchung des” Herzens “einer kleinen späten Kreidezeit Thescelosaurus“. Naturwissenschaften. 98 (3): 203–211. Bibcode:2011NW ….. 98..203C. doi:10.1007 / s00114-010-0760-1. PMID 21279321. S2CID 2408562.CS1-Wartung: mehrere Namen: Autorenliste (Link)
- ^ Chinsamy, Anusuya; und Hillenius, Willem J. (2004). “Physiologie nichtavianischer Dinosaurier”. Die Dinosaurier, 2 .. 643–659.
- ^ Xu, X.; Norell, MA (2004). “Ein neuer troodontischer Dinosaurier aus China mit einer vogelähnlichen Schlafhaltung”. Natur. 431 (7010): 838–41. Bibcode:2004Natur.431..838X. doi:10.1038 / nature02898. PMID 15483610. S2CID 4362745.Siehe Kommentar zum Artikel
- ^ Schweitzer, MH; Wittmeyer, JL; und Horner, JR (2005). “Geschlechtsspezifisches Fortpflanzungsgewebe bei Laufvögeln und Tyrannosaurus Rex“. Wissenschaft. 308 (5727): 1456–60. Bibcode:2005Sci … 308.1456S. doi:10.1126 / science.1112158. PMID 15933198. S2CID 30264554.CS1-Wartung: mehrere Namen: Autorenliste (Link)
- ^ Lee, Andrew H.; Werning, Sarah (2008). “Geschlechtsreife bei wachsenden Dinosauriern passt nicht zu Reptilienwachstumsmodellen”. Verfahren der Nationalen Akademie der Wissenschaften. 105 (2): 582–7. Bibcode:2008PNAS..105..582L. doi:10.1073 / pnas.0708903105. PMC 2206579. PMID 18195356.
- ^ MA Norell, JM Clark, D. Dashzeveg, T. Barsbold, LM Chiappe, AR Davidson, MC McKenna und MJ Novacek (November 1994). “Ein Theropod-Dinosaurier-Embryo und die Affinitäten der Flaming Cliffs Dinosaurier-Eier”. Wissenschaft. 266 (5186): 779–82. Bibcode:1994Sci … 266..779N. doi:10.1126 / science.266.5186.779. PMID 17730398. S2CID 22333224.CS1-Wartung: mehrere Namen: Autorenliste (Link)
- ^ Wings O (2007). “Eine Überprüfung der Gastrolithfunktion mit Auswirkungen auf fossile Wirbeltiere und eine überarbeitete Klassifizierung”. Palaeontologica Polonica. 52 (1): 1–16.
- ^ Embery, G; Milner, AC; Waddington, RJ; Hall, RC; Langley, MS; Milan, AM (2003). “Identifizierung von proteinhaltigem Material im Knochen des Dinosauriers Iguanodon”. Bindegewebsforschung. 44 (Suppl 1): 41–6. doi:10.1080 / 03008200390152070. PMID 12952172. S2CID 2249126.
- ^ Schweitzer, MH; Marshall, M; Carron, K; Bohle, DS; Busse, SC; Arnold, EV; Barnard, D; Horner, JR; Starkey, JR (Juni 1997). “Hämverbindungen im Trabekelknochen von Dinosauriern”. Verfahren der National Academy of Sciences der Vereinigten Staaten von Amerika. 94 (12): 6291–6. Bibcode:1997PNAS … 94.6291S. doi:10.1073 / pnas.94.12.6291. PMC 21042. PMID 9177210.
- ^ Schweitzer, MH; Wittmeyer, JL; Horner, JR; Toporski, JK (2005). “Weichteilgefäße und Zellkonservierung in Tyrannosaurus rex”. Wissenschaft. 307 (5717): 1952–5. Bibcode:2005Sci … 307.1952S. doi:10.1126 / science.1108397. PMID 15790853. S2CID 30456613. Behandelt auch den Abschnitt “Reproduktionsbiologie” im Abschnitt “Gefiederte Dinosaurier” und “Vogelverbindung”.
- ^ Asara, JM; Schweitzer MH, Freimark LM, Phillips M., Cantley LC (2007). “Proteinsequenzen von Mastodon und Tyrannosaurus Rex durch Massenspektrometrie enthüllt”. Wissenschaft. 316 (5822): 280–5. Bibcode:2007Sci … 316..280A. doi:10.1126 / science.1137614. PMID 17431180. S2CID 85299309.CS1-Wartung: mehrere Namen: Autorenliste (Link)
- ^ Orgel, CL; Schweitzer, MH; Zheng, W; Freimark, LM; Cantley, LC; Asara, JM (2008). “Molekulare Phylogenetik von Mastodon und Tyrannosaurus Rex“. Wissenschaft. 320 (5875): 499. doi:10.1126 / science.1154284. PMID 18436782. S2CID 24971064.
- ^ Schweitzer, MH; Zheng W., Orgel CL, Avci R., Suo Z., Freimark LM, Lebleu VS, Duncan MB, Vander Heiden MG, Neveu JM, Lane WS, Cottrell JS, Horner JR, Cantley LC, Kalluri R. und Asara JM ( 2009). “Biomolekulare Charakterisierung und Proteinsequenzen des kampanischen Hadrosauriers B. canadensis”. Wissenschaft. 324 (5927): 626–31. Bibcode:2009Sci … 324..626S. doi:10.1126 / science.1165069. PMID 19407199. S2CID 5358680.CS1-Wartung: mehrere Namen: Autorenliste (Link)
- ^ Kaye, TG; Gaugler, G; Sawlowicz, Z (2008). “Dinosaurier-Weichteile als bakterielle Biofilme interpretiert”. PLUS EINS. 3 (7): e2808. Bibcode:2008PLoSO … 3.2808K. doi:10.1371 / journal.pone.0002808. PMC 2483347. PMID 18665236.
- ^ Peterson, JE; Lenczewski, ME; Scherer, RP (2010). “Einfluss mikrobieller Biofilme auf die Erhaltung des primären Weichgewebes bei fossilen und vorhandenen Archosauriern”. PLUS EINS. 5 (10): e13334. Bibcode:2010PLoSO … 513334P. doi:10.1371 / journal.pone.0013334. PMC 2953520. PMID 20967227.
- ^ Bern, M; Phinney, BS; Goldberg, D (2009). “Reanalyse von Tyrannosaurus Rex Massenspektren “. Journal of Proteome Research. 8 (9): 4328–4332. doi:10.1021 / pr900349r. PMC 2738754. PMID 19603827.
- ^ Cleland, TP; et al. (2015). Massenspektrometrie und antikörperbasierte Charakterisierung von Blutgefäßen aus Brachylophosaurus canadensis“”. Journal of Proteome Research. 14 (12): 5252–5262. doi:10.1021 / acs.jproteome.5b00675. PMC 4768904. PMID 26595531.
- ^ Buckley, Michael; Warwood, Stacey; van Dongen, Bart; Kitchener, Andrew C.; Manning, Phillip L. (2017). “Eine fossile Protein-Chimäre; Schwierigkeiten bei der Unterscheidung von Dinosaurier-Peptidsequenzen von moderner Kreuzkontamination”. Verfahren der Royal Society B.. 284 (1855): 20170544. doi:10.1098 / rspb.2017.0544. PMC 5454271. PMID 28566488.
- ^ Wang, HL; Yan, ZY; Jin, DY (1997). “Reanalyse der veröffentlichten DNA-Sequenz, die aus Fossilien von Kreide-Dinosaurier-Eiern amplifiziert wurde”. Molekularbiologie und Evolution. 14 (5): 589–91. doi:10.1093 / oxfordjournals.molbev.a025796. PMID 9159936.
- ^ Voeten, Dennis FAE; et al. (13. März 2018). “Flügelknochengeometrie zeigt aktiven Flug in Archaeopteryx”. Naturkommunikation. 9 (923): 923. Bibcode:2018NatCo … 9..923V. doi:10.1038 / s41467-018-03296-8. PMC 5849612. PMID 29535376.
- ^ Guarino, Ben (13. März 2018). “Dieser gefiederte Dinosaurier ist wahrscheinlich geflogen, aber nicht wie jeder Vogel, den du kennst.”. Die Washington Post. Abgerufen 13. März 2018.
- ^ Terres, John K. (1980). Die Audubon Society Encyclopedia of North American Birds. New York, NY: Knopf. pp. 398–401. ISBN 978-0-394-46651-4.
- ^ Poling, J. (1996). “Federn, Rillen und der Ursprung der Vögel”. dinosauria.com. Archiviert von das Original am 2008-04-02. Abgerufen 2008-03-21.
- ^ Prum, R. & Brush, AH (2002). “Der evolutionäre Ursprung und die Diversifizierung von Federn” (PDF). Die vierteljährliche Überprüfung der Biologie. 77 (3): 261–95. doi:10.1086 / 341993. PMID 12365352. S2CID 6344830. Archiviert von das Original (PDF) am 15.10.2003. Abgerufen 2018-09-17.
- ^ Mayr G., Pohl B., Peters D. (2005). “Eine gut erhaltene Archaeopteryx Probe mit Theropodenmerkmalen “. Wissenschaft. 310 (5753): 1483–6. Bibcode:2005Sci … 310.1483M. doi:10.1126 / science.1120331. PMID 16322455. S2CID 28611454.
- ^ Feduccia, A. (1999). Der Ursprung und die Entwicklung der Vögel. Yale University Press. ISBN 978-0-300-07861-9.
- ^ Feduccia, A. (Februar 1995). “Explosive Evolution bei tertiären Vögeln und Säugetieren”. Wissenschaft. 267 (5198): 637–8. Bibcode:1995Sci … 267..637F. doi:10.1126 / science.267.5198.637. PMID 17745839. S2CID 42829066. Archiviert von das Original am 20.03.2008.
- ^ Feduccia, A. (1993).[specify]
- ^ Es wurden Kreidespuren eines Vogels mit einem ähnlichen Lebensstil gefunden – Lockley, MG, Li, R., Harris, JD, Matsukawa, M. und Liu, M. (August 2007). “Früheste Zygodactyl-Vogelfüße: Beweise aus Roadrunner-ähnlichen Spuren der frühen Kreidezeit”. Naturwissenschaften. 94 (8): 657–65. Bibcode:2007NW ….. 94..657L. doi:10.1007 / s00114-007-0239-x. PMID 17387416. S2CID 15821251.CS1-Wartung: mehrere Namen: Autorenliste (Link)
- ^ Burgers, P. & LM Chiappe (1999). “Der Flügel des Archaeopteryx als primärer Schubgenerator”. Natur. 399 (6731): 60–62. Bibcode:1999Natur.399 … 60B. doi:10.1038 / 19967. S2CID 4430686.
- ^ Cowen, R. Geschichte des Lebens. Blackwell Science. ISBN 978-0-7266-0287-0.
- ^ Videler, JJ 2005: Vogelflug. Universität Oxford. Drücken Sie, Oxford.
- ^ Burke, AC & amp; Feduccia, A. (1997). “Entwicklungsmuster und die Identifizierung von Homologien in der Vogelhand”. Wissenschaft. 278 (5338): 666–668. Bibcode:1997Sci … 278..666B. doi:10.1126 / science.278.5338.666. Zusammengefasst bei “Embryo-Studien zeigen, dass Dinosaurier modernen Vögeln keinen Aufstieg geben konnten”. ScienceDaily. Oktober 1997.
- ^ Chatterjee, S. (April 1998). “Die Finger von Vögeln und Dinosauriern zählen”. Wissenschaft. 280 (5362): 355a – 355. Bibcode:1998Sci … 280..355C. doi:10.1126 / science.280.5362.355a.
- ^ Vargas, AO, Fallon, JF (Oktober 2004). “Vögel haben Dinosaurierflügel: Der molekulare Beweis”. Journal of Experimental Zoology Teil B: Molekulare und Entwicklungsentwicklung. 304B (1): 86–90. doi:10.1002 / jez.b.21023. PMID 15515040.CS1-Wartung: mehrere Namen: Autorenliste (Link)
- ^ Pennisi, E. (Januar 2005). “Vogelflügel sind wirklich wie Dinosaurierhände” (PDF). Wissenschaft. 307 (5707): 194b. doi:10.1126 / science.307.5707.194b. PMID 15653478. S2CID 82490156. Archiviert von das Original (PDF) am 27.07.2011.
- ^ Es gibt einen Videoclip eines sehr jungen Mädchens, das dies bei tut “Flügelunterstütztes Steigungslaufen und Flugentwicklung”.
- ^ Dial, KP (2003). “Wing-Assisted Incline Running und die Entwicklung des Fluges”. Wissenschaft. 299 (5605): 402–4. Bibcode:2003Sci … 299..402D. doi:10.1126 / science.1078237. PMID 12532020. S2CID 40712093. Zusammengefasst in Morelle, Rebecca (24. Januar 2008). “Geheimnisse des Vogelfluges enthüllt” (Netz). Wissenschaftler glauben, dass sie der Lösung des Rätsels, wie die ersten Vögel in die Luft gingen, einen Schritt näher kommen könnten. BBC News. Abgerufen 2008-01-25.
- ^ Bundle, MW & Dial, KP (2003). “Mechanik des flügelunterstützten Steigungslaufs (WAIR)” (PDF). Das Journal of Experimental Biology. 206 (Pt 24): 4553–64. doi:10.1242 / jeb.00673. PMID 14610039. S2CID 6323207.
- ^ ein b Senter, P. (2006). “Skapulierorientierung bei Theropoden und Basalvögeln und der Ursprung des Schlagflugs”. Acta Palaeontologica Polonica. 51 (2): 305–313.
- ^
- ^ Chatterjee, Sankar, Templin, RJ (2004) “Gefiederte Coelurosaurier aus China: Neues Licht auf den baumartigen Ursprung des Vogelfluges”, S. 251-281. In Feathered Dragons: Studien zum Übergang von Dinosauriern zu Vögeln (PJ Currie, EB Koppelhus, MA Shugar und JL Wright (Hrsg.). Indiana University Press, Bloomington.
- ^ Tarsitano, Samuel F.; Russell, Anthony P.; Horne, Francis; Stehlager, Christopher; Millerchip, Karen (2000). “Über die Entwicklung der Federn aus aerodynamischer und konstruktiver Sicht”. Amerikanischer Zoologe. 40 (4): 676–686. doi:10.1093 / icb / 40.4.676.
- ^ Hu, D.; Hou, L.; Zhang, L. & Xu, X. (2009). “Ein Pre-Archaeopteryx Troodontid Theropod aus China mit langen Federn am Mittelfuß “. Natur. 461 (7264): 640–3. Bibcode:2009Natur.461..640H. doi:10.1038 / nature08322. PMID 19794491. S2CID 205218015.
- ^ Hopson, James A. “Ökomorphologie der phalangealen Proportionen von aviären und nichtavianischen Theropoden: Implikationen für den arborealen versus terrestrischen Ursprung des Vogelfluges” (2001) Aus neuen Perspektiven auf den Ursprung und die frühe Entwicklung von Vögeln: Proceedings of the International Symposium zu Ehren von John H. Ostrom. J. Gauthier und LF Gall, Hrsg. New Haven: Peabody Mus. Nat. Hist., Yale Univ. ISBN 0-912532-57-2. © 2001 Peabody Museum of Natural History, Yale University. Alle Rechte vorbehalten.
- ^ Glen, CL & Bennett, MB (November 2007). “Futtersuchmodi von mesozoischen Vögeln und Nicht-Vogel-Theropoden”. Aktuelle Biologie. 17 (21): R911–2. doi:10.1016 / j.cub.2007.09.026. PMID 17983564. S2CID 535424. Archiviert von das Original am 08.12.2012.
- ^ Alonso, PD, Milner, AC, Ketcham, RA, Cokson, MJ und Rowe, TB (August 2004). “Die Vogelart des Gehirns und des Innenohrs von Archaeopteryx”. Natur. 430 (7000): 666–9. Bibcode:2004Natur.430..666A. doi:10.1038 / nature02706. PMID 15295597. S2CID 4391019.CS1-Wartung: mehrere Namen: Autorenliste (Link)
- ^ Chiappe, LM (2007-02-02). Verherrlichte Dinosaurier: Der Ursprung und die frühe Entwicklung der Vögel. Sydney: UNSW Press. ISBN 978-0-471-24723-4.
- ^ Zhang, F., Zhou, Z., Xu, X. & Wang, X. (2002). “Ein jugendlicher Coelurosaurier-Theropod aus China weist auf Baumgewohnheiten hin”. Naturwissenschaften. 89 (9): 394–8. Bibcode:2002NW ….. 89..394Z. doi:10.1007 / s00114-002-0353-8. PMID 12435090. S2CID 556221.CS1-Wartung: mehrere Namen: Autorenliste (Link)
- ^ Chatterjee, S; Templin, RJ (2007). “Doppeldecker-Flügelplanform und Flugleistung des gefiederten Dinosauriers Microraptor gui” (PDF). Verfahren der Nationalen Akademie der Wissenschaften. 104 (5): 1576–80. Bibcode:2007PNAS..104.1576C. doi:10.1073 / pnas.0609975104. PMC 1780066. PMID 17242354.
- ^ Beebe, CWA (1915). “Tetrapteryx-Stadium in der Abstammung der Vögel”. Zoologica. 2: 38–52.
- ^ Lucas, Frederic A. (Frederic Augustus); Amerikanisches Naturkundemuseum (13. Juni 2018). Tiere der Vergangenheit: Ein Bericht über einige der Kreaturen der Antike. New York : [Amer. Mus. Press] – über das Internetarchiv.
- ^ Paul, GS (2002). “Dinosaurier der Luft: Die Entwicklung und der Verlust des Fluges bei Dinosauriern und Vögeln.” Baltimore: Johns Hopkins University Press. Seite 257
- ^ Corfe, IJ; Butler, RJ (2006). “Kommentieren Eine gut erhaltene Archaeopteryx-Probe mit Theropod-Merkmalen“. Wissenschaft. 313 (5791): 1238b. Bibcode:2006Sci … 313.1238C. doi:10.1126 / science.1130800. PMID 16946054. S2CID 19859320.
- ^ Parsons, William L.; Parsons, Kristen M. (2015). Morphologische Variationen innerhalb der Ontogenese von Deinonychus antirrhopus (Theropoda, Dromaeosauridae). PLUS EINS. 10 (4): e0121476. Bibcode:2015PLoSO..1021476P. doi:10.1371 / journal.pone.0121476. PMC 4398413. PMID 25875499.
- ^ Hutson, Joel D.; Hutson, Kelda N. (2018). “Die Beibehaltung des flugangepassten Vogel-Finger-Gelenk-Komplexes im Strauß hilft zu erkennen, wann sich Flügel bei Dinosauriern zu entwickeln begannen.” Strauß: Journal of African Ornithology. 89 (2): 173–186. doi:10.2989 / 00306525.2017.1422566. S2CID 91081348.
- ^ Chiappe, Luis (2002-01-01), Osteologie des flugunfähigen Patagopteryx deferrariisi aus der späten Kreidezeit Patagoniens (Argentinien)S. 281–316, ISBN 978-0-520-20094-4abgerufen 2020-07-27
Verweise[edit]
- Barsbold, Rinchen (1983): O ptich’ikh chertakh gegen stroyenii khishchnykh dinozavrov. [“Avian” features in the morphology of predatory dinosaurs]. Transaktionen der gemeinsamen sowjetischen mongolischen paläontologischen Expedition 24: 96-103. [Original article in Russian.] Übersetzt von W. Robert Welsh, Kopie von Kenneth Carpenter und konvertiert von Matthew Carrano. PDF Volltext
- Borenstein, Seth (31. Juli 2014). “Studie verfolgt die Entwicklung der Dinosaurier zu Frühaufstehern”. AP Nachrichten. Abgerufen 3. August 2014.
- Bostwick, Kimberly S. (2003). “Ursprung und Entwicklung der Vögel: Daten sammeln sich, Wissenschaftler integrieren sich, und dennoch tobt die” Debatte “. Kladistik. 19 (4): 369–371. doi:10.1016 / S0748-3007 (03) 00069-0.
- Dingus, Lowell & Rowe, Timothy (1997): Das irrtümliche Aussterben: Dinosaurier-Evolution und der Ursprung der Vögel. WH Freeman and Company, New York. ISBN 0-7167-2944-X
- Dinosauria Online (1995): Archaeopteryx ‘Beziehung zu modernen Vögeln. Abgerufen am 30.09.2006.
- Dinosauria Online (1996): Dinosaurier-Synapomorphien gefunden in Archaeopteryx. Abgerufen am 30.09.2006.
- Heilmann, G. (1926): Der Ursprung der Vögel. Witherby, London. ISBN 0-486-22784-7 (1972 Dover Nachdruck)
- Mayr, Gerald; Pohl, B.; Peters, DS (2005). “Ein gut erhaltener Archaeopteryx Probe mit Theropod-Merkmalen “. Wissenschaft. 310 (5753): 1483–1486. Bibcode:2005Sci … 310.1483M. doi:10.1126 / science.1120331. PMID 16322455. S2CID 28611454.
- Olson, Storrs L. (1985): Der Fossilienbestand von Vögeln. Im: Farner, DS; King, JR & Parkes, Kenneth C. (Hrsg.): Vogelbiologie 8: 79-238. Akademische Presse, New York.
Externe Links[edit]
Recent Comments