Meselson-Stahl-Experiment – Wikipedia
Die Meselson-Stahl-Experiment ist ein Experiment von Matthew Meselson und Franklin Stahl aus dem Jahr 1958, das Watson und Cricks Hypothese stützte, dass die DNA-Replikation semikonservativ war. Bei der semikonservativen Replikation, wenn die doppelsträngige DNA-Helix repliziert wird, bestand jede der beiden neuen doppelsträngigen DNA-Helices aus einem Strang der ursprünglichen Helix und einem neu synthetisierten. Es wurde “das schönste Experiment der Biologie” genannt.[1] Meselson und Stahl entschieden, dass der beste Weg, die Eltern-DNA zu markieren, darin besteht, eines der Atome im Eltern-DNA-Molekül zu ändern. Da Stickstoff in den stickstoffhaltigen Basen jedes Nukleotids gefunden wird, entschieden sie sich, ein Stickstoffisotop zu verwenden, um zwischen Eltern- und neu kopierter DNA zu unterscheiden. Das Stickstoffisotop hatte ein zusätzliches Neutron im Kern, was es schwerer machte.
Hypothese[edit]
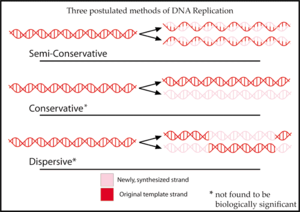
Drei Hypothesen wurden zuvor für das Verfahren der DNA-Replikation vorgeschlagen.
In dem halbkonservativ Hypothese, vorgeschlagen von Watson und Crick, trennen sich die beiden Stränge eines DNA-Moleküls während der Replikation. Jeder Strang fungiert dann als Matrize für die Synthese eines neuen Strangs.[2]
Die konservativ Die Hypothese schlug vor, dass das gesamte DNA-Molekül als Matrize für die Synthese eines völlig neuen Moleküls diente. Nach diesem Modell binden Histonproteine an die DNA, drehen den Strang und setzen die Nukleotidbasen (die normalerweise das Innere auskleiden) für Wasserstoffbrücken frei.[3]
Die dispersiv Die Hypothese wird durch ein von Max Delbrück vorgeschlagenes Modell veranschaulicht, das versucht, das Problem der Entwindung der beiden Stränge der Doppelhelix durch einen Mechanismus zu lösen, der das DNA-Rückgrat etwa alle 10 Nukleotide bricht, das Molekül aufdreht und den alten Strang an das Ende des neu synthetisierten. Dies würde die DNA in kurzen Stücken synthetisieren, die sich von einem Strang zum anderen abwechseln.[4]
Jedes dieser drei Modelle macht eine andere Vorhersage über die Verteilung der „alten“ DNA in Molekülen, die nach der Replikation gebildet werden. In der konservativen Hypothese ist nach der Replikation ein Molekül das vollständig konservierte “alte” Molekül und das andere ist komplett neu synthetisierte DNA. Die semikonservative Hypothese sagt voraus, dass jedes Molekül nach der Replikation einen alten und einen neuen Strang enthält. Das dispersive Modell sagt voraus, dass jeder Strang jedes neuen Moleküls eine Mischung aus alter und neuer DNA enthält.[5]
Versuchsdurchführung und Ergebnisse[edit]
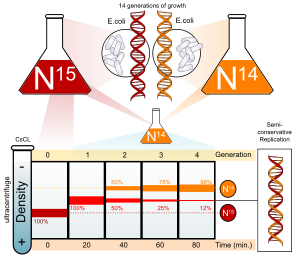
Stickstoff ist ein Hauptbestandteil der DNA. 14N ist bei weitem das häufigste Isotop von Stickstoff, aber DNA mit dem schwereren (aber nicht radioaktiven) fünfzehnN-Isotop ist auch funktionell.
E coli wurde über mehrere Generationen in einem NH .-haltigen Medium gezüchtet4Cl mit fünfzehnN. Wenn DNA aus diesen Zellen extrahiert und auf einem Salz-(CsCl)-Dichtegradienten zentrifugiert wird, trennt sich die DNA an dem Punkt ab, an dem ihre Dichte der der Salzlösung entspricht. Die DNA der Zellen, die in fünfzehnN-Medium hatte eine höhere Dichte als Zellen, die in normalen 14N mittel. Danach, E coli Zellen mit nur fünfzehnN in ihrer DNA wurden auf a . übertragen 14N-Medium und durften sich teilen; der Fortschritt der Zellteilung wurde durch mikroskopische Zellzählungen und durch Kolonieassays überwacht.
DNA wurde periodisch extrahiert und mit reinem verglichen 14N DNA und fünfzehnN-DNA. Nach einer Replikation wurde gefunden, dass die DNA eine mittlere Dichte aufwies. Da eine konservative Replikation zu gleichen Mengen an DNA mit höherer und niedrigerer Dichte führen würde (aber keine DNA mit mittlerer Dichte), wurde eine konservative Replikation ausgeschlossen. Dieses Ergebnis stimmte jedoch sowohl mit der semikonservativen als auch mit der dispersiven Replikation überein. Semikonservative Replikation würde zu doppelsträngiger DNA mit einem Strang von fünfzehnN DNA und einer von 14N-DNA, während dispersive Replikation zu doppelsträngiger DNA führen würde, wobei beide Stränge Mischungen von fünfzehnN und 14N-DNA, von denen jede als DNA mittlerer Dichte erschienen wäre.
Die Autoren fuhren fort, Zellen zu beproben, während die Replikation fortgesetzt wurde. Es wurde festgestellt, dass DNA aus Zellen nach Abschluss zweier Replikationen aus gleichen Mengen DNA mit zwei unterschiedlichen Dichten besteht, von denen eine der mittleren DNA-Dichte von Zellen entspricht, die für nur eine Teilung in gezüchtet wurden 14N-Medium, das andere entspricht DNA aus Zellen, die ausschließlich in . gezüchtet wurden 14N mittel. Dies war unvereinbar mit der dispersiven Replikation, die zu einer einzigen Dichte geführt hätte, die niedriger als die Zwischendichte der Zellen einer Generation war, aber immer noch höher als bei Zellen, die nur in . gezüchtet wurden 14N DNA-Medium, wie das Original fünfzehnN DNA wäre gleichmäßig auf alle DNA-Stränge aufgeteilt worden. Das Ergebnis stimmte mit der semikonservativen Replikationshypothese überein.[6]
Verweise[edit]
- ^ John Cairns an Horace F. Judson, in The Eighth Day of Creation: Makers of the Revolution in Biology (1979). Touchstone-Bücher, ISBN 0-671-22540-5. 2. Auflage: Cold Spring Harbor Laboratory Press, 1996 Taschenbuch: ISBN 0-87969-478-5.
- ^ Watson JD, Crick FH (1953). „Die Struktur der DNA“. Kalter Frühlingshafen. Symp. Menge Biol. 18: 123–31. mach:10.1101/SQB.1953.018.01.020. PMID 13168976.
- ^ Bloch DP (Dezember 1955). “Ein möglicher Mechanismus für die Replikation der helikalen Struktur von Desoxyribonukleinsäure”. Proz. Natl. Akad. Wissenschaft Vereinigte Staaten von Amerika 41 (12): 1058–64. Bibcode:1955PNAS…41.1058B. mach:10.1073/pnas.41.12.1058. PMC 528197. PMID 16589796.
- ^ Delbrück M (September 1954). “Über die Replikation von Desoxyribonukleinsäure (DNA)” (PDF). Proz. Natl. Akad. Wissenschaft Vereinigte Staaten von Amerika 40 (9): 783–8. Bibcode:1954PNAS…40..783D. mach:10.1073/pnas.40.9.783. PMC 534166. PMID 16589559.
- ^ Delbrück, Max; Stent, Gunther S. (1957). „Über den Mechanismus der DNA-Replikation“. In McElroy, William D.; Glas, Bentley (Hrsg.). Ein Symposium über die chemischen Grundlagen der Vererbung. Johns HopkinsPr. pp. 699-736.
- ^ Meselson, M. & Stahl, FW (1958). “Die Replikation der DNA in Escherichia coli”. PNAS. 44 (7): 671–82. Bibcode:1958PNAS…44..671M. mach:10.1073/pnas.44.7.671. PMC 528642. PMID 16590258.
Externe Links[edit]
Recent Comments