Sauerstoff-Hämoglobin-Dissoziationskurve – Wikipedia
Die Sauerstoff-Hämoglobin-Dissoziationskurve, auch genannt Oxyhämoglobin-Dissoziationskurve oder Sauerstoffdissoziationskurve (ODC), ist eine Kurve, die den Anteil von Hämoglobin in seiner gesättigten (sauerstoffbeladenen) Form auf der vertikalen Achse gegen die vorherrschende Sauerstoffspannung auf der horizontalen Achse aufträgt. Diese Kurve ist ein wichtiges Werkzeug, um zu verstehen, wie unser Blut Sauerstoff transportiert und freisetzt. Insbesondere bezieht sich die Oxyhämoglobin-Dissoziationskurve auf die Sauerstoffsättigung (SÖ2) und Sauerstoffpartialdruck im Blut (PÖ2) und wird durch die sogenannte “Hämoglobin-Affinität für Sauerstoff” bestimmt; das heißt, wie leicht Hämoglobin Sauerstoffmoleküle aufnimmt und in die es umgebende Flüssigkeit abgibt.

Hintergrund[edit]
Hämoglobin (Hb) ist das wichtigste Vehikel für den Sauerstofftransport im Blut. Jedes Hämoglobinmolekül hat die Kapazität, vier Sauerstoffmoleküle zu tragen. Diese Sauerstoffmoleküle binden an das Eisen der Hämprothesengruppe.[1]
Wenn Hämoglobin weder gebundenen Sauerstoff noch gebundenes Kohlendioxid hat, hat es die ungebundene Konformation (Form). Die Bindung des ersten Sauerstoffmoleküls führt zu einer Formänderung des Hämoglobins, die seine Fähigkeit zur Bindung an die anderen drei Sauerstoffmoleküle erhöht.
In Gegenwart von gelöstem Kohlendioxid ändert sich der pH-Wert des Blutes; Dies führt zu einer weiteren Formänderung des Hämoglobins, die seine Fähigkeit zur Bindung von Kohlendioxid erhöht und seine Fähigkeit zur Bindung von Sauerstoff verringert. Mit dem Verlust des ersten Sauerstoffmoleküls und der Bindung des ersten Kohlendioxidmoleküls tritt noch eine weitere Formänderung auf, die die Fähigkeit zur Sauerstoffbindung weiter verringert und die Fähigkeit zur Kohlendioxidbindung erhöht. Der an das Hämoglobin gebundene Sauerstoff wird an das Blutplasma abgegeben und vom Gewebe absorbiert, und das Kohlendioxid im Gewebe wird an das Hämoglobin gebunden.
In der Lunge findet der umgekehrte Vorgang statt. Mit dem Verlust des ersten Kohlendioxidmoleküls ändert sich die Form wieder und erleichtert die Freisetzung der anderen drei Kohlendioxide.
Sauerstoff wird auch im Blutplasma gelöst transportiert, jedoch in viel geringerem Maße. Hämoglobin ist in roten Blutkörperchen enthalten. Hämoglobin setzt den gebundenen Sauerstoff frei, wenn Kohlensäure vorhanden ist, wie sie im Gewebe vorhanden ist. In den Kapillaren, in denen Kohlendioxid produziert wird, wird an das Hämoglobin gebundener Sauerstoff in das Blutplasma abgegeben und in das Gewebe aufgenommen.
Wie viel dieser Kapazität zu einem bestimmten Zeitpunkt mit Sauerstoff gefüllt ist, wird als Sauerstoffsättigung bezeichnet. In Prozent ausgedrückt ist die Sauerstoffsättigung das Verhältnis der an das Hämoglobin gebundenen Sauerstoffmenge zur Sauerstofftragfähigkeit des Hämoglobins. Die Sauerstofftransportkapazität von Hämoglobin wird durch die Art des im Blut vorhandenen Hämoglobins bestimmt. Die zu jeder Zeit an das Hämoglobin gebundene Sauerstoffmenge hängt zum großen Teil vom Sauerstoffpartialdruck ab, dem das Hämoglobin ausgesetzt ist. In der Lunge ist der Sauerstoffpartialdruck an der Alveolar-Kapillar-Grenzfläche typischerweise hoch, und daher bindet der Sauerstoff leicht an vorhandenes Hämoglobin. Wenn das Blut zu anderen Körpergeweben zirkuliert, in denen der Sauerstoffpartialdruck geringer ist, gibt das Hämoglobin den Sauerstoff in das Gewebe ab, da das Hämoglobin in Gegenwart niedrigerer Sauerstoffpartialdrücke seine volle Sauerstoffbindungskapazität nicht aufrechterhalten kann.
Sigmaform[edit]

Die Kurve wird normalerweise am besten durch ein Sigmoid-Plot beschrieben, wobei eine Formel der Art verwendet wird:
-
Die Stärke, mit der Sauerstoff an Hämoglobin bindet, wird von mehreren Faktoren beeinflusst. Diese Faktoren verschieben oder formen die Oxyhämoglobin-Dissoziationskurve um. Eine Rechtsverschiebung zeigt an, dass das untersuchte Hämoglobin eine verringerte Affinität zu Sauerstoff hat. Dies erschwert die Bindung des Hämoglobins an Sauerstoff (erfordert einen höheren Sauerstoffpartialdruck, um die gleiche Sauerstoffsättigung zu erreichen), aber es erleichtert dem Hämoglobin die Freisetzung von daran gebundenem Sauerstoff. Die Wirkung dieser Rechtsverschiebung der Kurve erhöht den Sauerstoffpartialdruck in den Geweben, wenn er am dringendsten benötigt wird, wie zum Beispiel während des Trainings oder eines hämorrhagischen Schocks. Im Gegensatz dazu wird die Kurve durch das Gegenteil dieser Bedingungen nach links verschoben. Diese Linksverschiebung weist darauf hin, dass das untersuchte Hämoglobin eine erhöhte Affinität zu Sauerstoff hat, so dass Hämoglobin Sauerstoff leichter bindet, ihn aber widerwilliger entlädt. Eine Linksverschiebung der Kurve ist ein Zeichen für die erhöhte Affinität des Hämoglobins zu Sauerstoff (zB an der Lunge). In ähnlicher Weise zeigt die Rechtsverschiebung eine verringerte Affinität, wie sie bei einer Erhöhung der Körpertemperatur, der Wasserstoffionen, der 2,3-Bisphosphoglycerat (2,3-BPG)-Konzentration oder der Kohlendioxidkonzentration erscheinen würde.
Kontrollfaktoren Ändern Kurvenverschiebung Temperatur ↑ → ↓ ← 2,3-BPG ↑ → ↓ ← pCO2 ↑ → ↓ ← Säure [H+] ↑ → ↓ ← Notiz:
- Linksverschiebung: höheres O2 Affinität
- Rechtsverschiebung: niedrigeres O2 Affinität
- fetales Hämoglobin hat einen höheren O2 Affinität als erwachsenes Hämoglobin; hauptsächlich aufgrund einer stark reduzierten Affinität zu 2,3-Bisphosphoglycerat.
Die Ursachen der Rechtsverschiebung können mit der Gedächtnisstütze erinnert werden, “KADETT, Gesicht Richtig!” für CÖ2, EINSäure, 2,3-DPG,[Note 1]Etrainieren und Temperatur.[2] Faktoren, die die Sauerstoffdissoziationskurve nach rechts verschieben, sind jene physiologischen Zustände, in denen Gewebe mehr Sauerstoff benötigen. Muskeln haben beispielsweise während des Trainings eine höhere Stoffwechselrate und benötigen folglich mehr Sauerstoff, produzieren mehr Kohlendioxid und Milchsäure und ihre Temperatur steigt.
pH[edit]
Eine Abnahme des pH-Wertes (Erhöhung der h+ Ionenkonzentration) verschiebt die Standardkurve nach rechts, eine Erhöhung nach links. Dies geschieht, weil bei größerem h+ Ionenkonzentration existieren verschiedene Aminosäurereste wie Histidin 146 überwiegend in ihrer protonierten Form, wodurch sie Ionenpaare bilden können, die Desoxyhämoglobin im T-Zustand stabilisieren.[3] Der T-Zustand hat eine geringere Affinität für Sauerstoff als der R-Zustand, daher bindet das Hämoglobin bei erhöhtem Säuregehalt weniger O2 für ein gegebenes PO2 (und mehr H+). Dies ist als Bohr-Effekt bekannt.[4] Eine Verringerung der Gesamtbindungskapazität von Hämoglobin an Sauerstoff (dh eine Verschiebung der Kurve nach unten, nicht nur nach rechts) aufgrund eines verringerten pH-Werts wird als Wurzeleffekt bezeichnet. Dies ist bei Knochenfischen zu sehen. Die Bindungsaffinität von Hämoglobin zu O2 ist bei einem relativ hohen pH-Wert am größten.
Kohlendioxid[edit]
Kohlendioxid beeinflusst die Kurve auf zwei Arten. Zuerst CO2 Akkumulation bewirkt die Bildung von Carbaminoverbindungen durch chemische Wechselwirkungen, die an Hämoglobin binden und Carbaminohämoglobin bilden. CO2 wird als allosterische Regulation angesehen, da die Hemmung nicht an der Bindungsstelle des Hämoglobins erfolgt.[5] Zweitens beeinflusst es den intrazellulären pH-Wert aufgrund der Bildung von Bicarbonationen. Die Bildung von Carbaminohämoglobin stabilisiert Hämoglobin im T-Zustand durch Bildung von Ionenpaaren.[3] Nur etwa 5–10 % des gesamten CO2 Der Blutgehalt wird als Carbamino-Verbindungen transportiert, während (80–90 %) als Bicarbonat-Ionen transportiert und eine kleine Menge im Plasma gelöst wird. Die Bildung eines Bicarbonat-Ions setzt ein Proton in das Plasma frei, wodurch der pH-Wert sinkt (erhöhter Säuregehalt), was auch die Kurve wie oben diskutiert nach rechts verschiebt; CO-arm2 Konzentrationen im Blutkreislauf führt zu einem hohen pH-Wert und bietet somit optimalere Bindungsbedingungen für Hämoglobin und O2. Dies ist ein physiologisch begünstigter Mechanismus, da Hämoglobin mehr Sauerstoff abgibt, wenn die Kohlendioxidkonzentration dramatisch ansteigt, wenn die Gewebeatmung schnell erfolgt und Sauerstoff benötigt wird.[6][7]
2,3-BPG[edit]
2,3-Bisphosphoglycerat oder 2,3-BPG (früher 2,3-Diphosphoglycerat oder 2,3-DPG – Referenz?) ist ein Organophosphat, das in roten Blutkörperchen während der Glykolyse gebildet wird und die konjugierte Base von 2,3-Bisphosphoglyceriden ist Säure. Die Produktion von 2,3-BPG ist wahrscheinlich ein wichtiger Anpassungsmechanismus, da die Produktion bei mehreren Erkrankungen in Gegenwart von vermindertem O . des peripheren Gewebes zunimmt2 Verfügbarkeit, wie unter anderem Hypoxämie, chronische Lungenerkrankung, Anämie und kongestive Herzinsuffizienz. Hohe 2,3-BPG-Werte verschieben die Kurve nach rechts (wie in der Kindheit), während niedrige 2,3-BPG-Werte eine Linksverschiebung verursachen, die bei Zuständen wie septischem Schock und Hypophosphatämie beobachtet wird.[4] In Abwesenheit von 2,3-BPG steigt die Affinität des Hämoglobins für Sauerstoff. 2,3-BPG wirkt als heteroallosterischer Effektor von Hämoglobin und senkt die Affinität des Hämoglobins für Sauerstoff, indem es vorzugsweise an Desoxyhämoglobin bindet. Eine erhöhte BPG-Konzentration in den roten Blutkörperchen begünstigt die Bildung des T (straff oder angespannt), des Hämoglobins mit niedriger Affinität, und so verschiebt sich die Sauerstoffbindungskurve nach rechts.
Temperatur[edit]
Eine Temperaturerhöhung verschiebt den ODC nach rechts. Wenn die Temperatur erhöht wird, halten Sie die
gleich, dann sinkt die Sauerstoffsättigung, weil die Bindung zwischen Eisen im und wird denaturiert. In ähnlicher Weise nimmt mit steigender Temperatur auch der Sauerstoffpartialdruck zu. Also hat man für das gleiche einen geringeren Hämoglobinsättigungsprozentsatz oder einen höheren Sauerstoffpartialdruck. Somit verschiebt sich jeder Punkt der Kurve nach rechts (aufgrund des erhöhten Sauerstoffpartialdrucks) und nach unten (aufgrund des abgeschwächten Bindung). Daher die Rechtsverschiebung der Kurve.[8]Kohlenmonoxid[edit]
Hämoglobin bindet sich 210-mal leichter an Kohlenmonoxid als an Sauerstoff.[4] Aufgrund dieser höheren Affinität von Hämoglobin zu Kohlenmonoxid als zu Sauerstoff ist Kohlenmonoxid ein sehr erfolgreicher Konkurrent, der selbst bei winzigen Partialdrücken Sauerstoff verdrängt. Die Reaktion HbO2 + CO → HbCO + O2 verdrängt fast irreversibel die Sauerstoffmoleküle, die das Carboxyhämoglobin bilden; die Bindung von Kohlenmonoxid an das Eisenzentrum von Hämoglobin ist viel stärker als die von Sauerstoff, und die Bindungsstelle bleibt für den Rest des Lebenszyklus des betroffenen roten Blutkörperchens blockiert.[9] Bei einem erhöhten Kohlenmonoxidspiegel kann eine Person an schwerer Gewebehypoxie leiden, während ein normaler pO .-Wert beibehalten wird2 weil Carboxyhämoglobin keinen Sauerstoff zu den Geweben transportiert.
Auswirkungen von Methämoglobinämie[edit]
Methämoglobinämie ist eine Form von abnormem Hämoglobin, bei der das Eisenzentrum vom Oxidationszustand Eisen(II) +2 (der normalen Form) in den Zustand von Eisen(III) +3 oxidiert wurde. Dies führt zu einer Linksverschiebung der Sauerstoffhämoglobin-Dissoziationskurve, da jedes Resthäm mit sauerstoffangereichertem Eisen(II) seinen gebundenen Sauerstoff nicht in das Gewebe entladen kann (weil 3+ Eisen die Kooperativität des Hämoglobins beeinträchtigt), wodurch seine Affinität zu Sauerstoff erhöht wird . Methämoglobin hat jedoch eine erhöhte Affinität zu Cyanid und ist daher bei der Behandlung von Cyanidvergiftungen nützlich. Im Falle einer versehentlichen Einnahme kann die Verabreichung eines Nitrits (wie Amylnitrit) verwendet werden, um das Hämoglobin absichtlich zu oxidieren und den Methämoglobinspiegel zu erhöhen, wodurch die Funktion der Cytochromoxidase wiederhergestellt wird. Das Nitrit wirkt auch als Vasodilatator, fördert die zelluläre Sauerstoffversorgung, und die Zugabe eines Eisensalzes sorgt für eine kompetitive Bindung des freien Cyanids als biochemisch inertes Hexacyanoferrat(III)-Ion. [Fe(CN)6]3−. Ein alternativer Ansatz beinhaltet die Verabreichung von Thiosulfat, wodurch Cyanid in Thiocyanat umgewandelt wird, SCN−, die ausgeschieden wird über die Nieren. Methämoglobin wird auch in geringen Mengen gebildet, wenn die Dissoziation von Oxyhämoglobin zur Bildung von Methämoglobin und Superoxid, O . führt2−, statt der üblichen Produkte. Superoxid ist ein freies Radikal und verursacht biochemische Schäden, wird jedoch durch die Wirkung des Enzyms Superoxid-Dismutase neutralisiert.
Auswirkungen von ITPP[edit]
Myo-Inositol-Trispyrophosphat (ITPP), auch bekannt als OXY111A, ist ein Inositol-Phosphat, das durch allosterische Modulation des Hämoglobins in den roten Blutkörperchen eine Rechtsverschiebung der Sauerstoff-Hämoglobin-Dissoziationskurve verursacht. Es ist ein experimentelles Medikament, das die Gewebehypoxie reduzieren soll. Die Wirkung scheint ungefähr so lange anzuhalten, wie die betroffenen roten Blutkörperchen im Kreislauf bleiben.
Fetales Hämoglobin[edit]
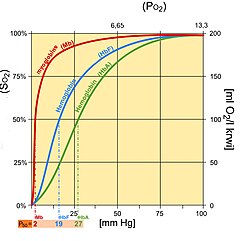 Sättigungskurve des fetalen Hämoglobins
Sättigungskurve des fetalen HämoglobinsFetales Hämoglobin (HbF) unterscheidet sich strukturell von normalem erwachsenem Hämoglobin (HbA), was HbF eine höhere Affinität für Sauerstoff verleiht als HbA. HbF besteht aus zwei Alpha- und zwei Gamma-Ketten, während HbA aus zwei Alpha- und zwei Beta-Ketten besteht. Aufgrund dieser strukturellen Unterschiede ist die fetale Dissoziationskurve gegenüber der Kurve für den normalen Erwachsenen nach links verschoben.
Typischerweise ist der arterielle Sauerstoffdruck des Fötus niedriger als der arterielle Sauerstoffdruck des Erwachsenen. Daher ist eine höhere Affinität zur Sauerstoffbindung bei niedrigeren Partialdruckniveaus im Fötus erforderlich, um die Diffusion von Sauerstoff durch die Plazenta zu ermöglichen. An der Plazenta wird eine höhere Konzentration von 2,3-BPG gebildet, und 2,3-BPG bindet eher an Beta-Ketten als an Alpha-Ketten. Als Ergebnis bindet 2,3-BPG stärker an adultes Hämoglobin, wodurch HbA mehr Sauerstoff zur Aufnahme durch den Fötus freisetzt, dessen HbF vom 2,3-BPG nicht beeinflusst wird.[10] HbF liefert dann den gebundenen Sauerstoff an Gewebe mit noch niedrigeren Partialdrücken, wo er freigesetzt werden kann.
Siehe auch[edit]
- ^ 2,3-DPG ist eine Abkürzung von 2,3-DichPhosphogLyzersäure, ein veralteter Name für 2,3-BPG
Verweise[edit]
- ^ Ahern, Kevin; Rajagopal, Indira; Tan, Taralyn (2017). Biochemie frei für alle (PDF) (1.2 Hrsg.). NC: Creative Commons.
- ^ “Medizinische Gedächtnisstütze”. LifeHugger. Abgerufen 2009-12-19.
- ^ ein B Lehninger. Prinzipien der Biochemie (6. Aufl.). P. 169.
- ^ ein B C Jacquez, John (1979). Atemphysiologie. McGraw-Hill. S. 156–175.
- ^ Ahern, Kevin; Rajagopal, Indira; Tan, Taralyn (5. August 2017). Biochemie frei für alle (1.2 Hrsg.). NC-Creative Commons. P. 370.
- ^ Ahern, Kevin; Rajagopal, Indira; Tan, Taralyn (5. August 2017). Biochemie frei für alle (1.2 Hrsg.). NC-Creative Commons. P. 134.
- ^ Donna, Larson (2017). Klinische Chemie: Grundlagen und Labortechniken. St. Louis, Missouri: Elsevier. P. 226. ISBN 978-1-4557-4214-1.
- ^ Schmidt-Nielsen (1997). Tierphysiologie: Anpassung und Umwelt. Cambridge University Press. ISBN 0521570980.
- ^ Kotz, John (August 2012). Chemie und chemische Reaktivität (8. Aufl.). Cengage-Lernen. P. 1032. ISBN 978-1133420071. Abgerufen 2015-07-01.
- ^ Lippincotts illustrierte Rezension: Biochemie 4. Auflage. Nordamerika: Lippincott, Williams und Wilkins. 2007. S. 24–35. ISBN 978-0-7817-6960-0.
Externe Links[edit]
![{displaystyle {ce {[O2]}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/8f8f06466530fdc7eb5967cec6cbfed6a59582aa "Sauerstoffkonzentration") gleich, dann sinkt die Sauerstoffsättigung, weil die Bindung zwischen Eisen im
gleich, dann sinkt die Sauerstoffsättigung, weil die Bindung zwischen Eisen im 


Recent Comments