Podokesaurus – Wikipedia
Coelophysoid Theropod Dinosaurier Gattung aus der frühen Jurazeit
Podokesaurus ist eine Gattung von Coelophysoid-Dinosauriern, die während der frühen Jurazeit im heutigen Osten der Vereinigten Staaten lebte. Das erste Fossil wurde 1910 vom Geologen Mignon Talbot in der Nähe von Mount Holyoke, Massachusetts, entdeckt. Das Exemplar war fragmentarisch und bewahrte einen Großteil des Körpers, der Gliedmaßen und des Schwanzes. 1911 beschrieb und benannte Talbot die neue Gattung und Art Podokesaurus Holyokensis basierend darauf. Der vollständige Name kann als “schnellfüßige Eidechse von Holyoke” übersetzt werden. Diese Entdeckung machte Talbot zur ersten Frau, die einen Dinosaurier fand und beschrieb. Das Holotyp-Fossil wurde als bedeutend erkannt und von anderen Forschern untersucht, ging jedoch verloren, als das Gebäude, in dem es aufbewahrt wurde, 1917 niedergebrannt wurde. keine eindeutige Podokesaurus Proben wurden seitdem entdeckt.
Geschätzt auf eine Länge von etwa 1 m und ein Gewicht von 1 bis 40 kg. Podokesaurus wurde leicht mit hohlen Knochen konstruiert und wäre ähnlich gewesen Coelophysis, schlank, langhalsig und mit scharfen, zurückgebogenen Zähnen. Das Wirbel waren sehr leicht und hohl, und einige waren an jedem Ende leicht konkav. Das zervikal (Hals-) Wirbel waren im Vergleich zu den relativ großen Längen und Durchmessern dorsal (Rücken-) Wirbel und die kaudal (Schwanz-) Wirbel waren lang und schlank. Das Humerus (Oberarmknochen) war klein und zart, weniger als die Hälfte der Länge des Femur (Oberschenkelknochen). Das Schambein war sehr lang und dehnte sich sowohl am vorderen als auch am hinteren Ende aus. Der Femur war schlank, fast gerade, hatte dünne Wände und war an der Rückseite seines unteren Endes erweitert. Die Drei Mittelfußknochen des Unterschenkels wurden eng zusammengedrückt und bildeten eine kompakte Struktur.
Da es sich um einen der wenigen kleinen Theropoden handelte, die zum Zeitpunkt der Beschreibung bekannt waren, waren die Affinitäten von Podokesaurus waren lange unklar. Es wurde zusammen mit anderen kleinen Theropoden in die Familie der Podokesauridae eingeordnet und soll einem Protovogel ähnlich gewesen sein. Es wurde vorgeschlagen, es sei ein Synonym für Coelophysis und ein natürliches gegossenes Exemplar wurde ihm zugewiesen, aber diese Ideen werden derzeit nicht akzeptiert. Die Familie Podokesauridae wird nicht mehr verwendet, da sie durch Coelophysidae ersetzt wurde, und Podokesaurus soll ein Coelophysoid gewesen sein. So wie, Podokesaurus wäre ein Raubtier mit leichtem Fuß gewesen, mit mächtigen Vorderbeinen und greifenden Händen. Es wird geschätzt, dass es mit 15 bis 20 km / h gelaufen sein könnte. Podokesaurus Es wird angenommen, dass es aus der Portland-Formation stammt, deren Alter lange unklar war, von der derzeit jedoch angenommen wird, dass sie vor 201 bis 190 Millionen Jahren auf die hettangisch-sinemurischen Stadien des frühen Jura datiert.
Entdeckung[edit]


1910 ging die amerikanische Geologin Mignon Talbot mit ihrer Schwester Ellen nach Holyoke, Massachusetts, im Osten der USA, als sie an einer Farm vorbeikamen und einen kleinen Hügel in der Nähe bemerkten. Es hatte eine Kiesgrube an einer Seite und wurde durch eine Ansammlung von Sand, Kies und Felsbrocken gebildet, die von einem zurückgehenden Gletscher zurückgelassen wurden. Talbot bemerkte einen weißen Streifen auf einem Sandsteinfelsen am Boden der Kiesgrube, und als sie entdeckte, dass es sich um Knochen handelte, sagte sie ihrer Schwester, sie habe ein “echtes lebendes Fossil” gefunden. Dies lag daran, dass zuvor im Connecticut Valley viele fossile Spuren entdeckt worden waren (zu denen sie ihre Schüler oft mitgenommen hatte), aber nur wenige echte Dinosaurierskelette und keine am Mount Holyoke. Der Landbesitzer erteilte ihr die Erlaubnis, das Exemplar für das Mount Holyoke College (ein Frauencollege ein paar Meilen von dort entfernt) zu sammeln, wo sie für die Abteilung für Geologie verantwortlich war.[1][2][3][4]
Am nächsten Tag brachte sie eine Gruppe von Arbeitern mit, um das Exemplar zu sammeln, und fand ein weiteres Stück Sandstein, das den Rest des Fossils sowie Eindrücke von denen in der ersten Platte enthielt. Das Exemplar schien jahrelang freigelegt zu sein, ohne dass es jemand bemerkte. Der Felsbrocken war von Menschen oder Frost aufgebrochen worden. Das Fossil wurde ins Labor gebracht, wo es vorbereitet und fotografiert wurde.[1] Das unvollständige Exemplar ist erhalten 5 zervikal (Hals), 11 dorsal (zurück) und 24 kaudal (Schwanz-) Wirbel, ein Fragment der linken Schulterblatt (Schulterblatt) und rechts Coracoid (Teil von dem Schultergürtel), ein Teil links Humerus (Oberarmknochen), Phalanxknochen von 3 Fingern, darunter 2 unguals (Klauenknochen), Rippen, die Scham und Ischium (Knochen, die die untere Vorder- bzw. untere Rückseite des Beckens bilden), die Femora (Oberschenkelknochen), links Tibia (Unterschenkelknochen), ein Fragment von rechts Astragalus (ein Knochen im Knöchel), artikuliert Mittelfußknochen (Fußknochen) des linken Fußes, gefolgt von 3 Pedal-Phalangen (Zehenknochen), partiellen Mittelfußknochen und möglichen Phalangen des rechten Fußes sowie Fragmenten unsicherer Identität.[5][6] Die leichten und empfindlichen Knochen befanden sich in ihrer natürlichen Position oder fast innerhalb des Felsens, mit Ausnahme des Schwanzes und der unsicheren Fragmente, die nach einer langen Lücke im Skelett hinter dem Becken einige Zentimeter vom Skelett entfernt waren. Die vordere Hälfte des Halses fehlte.[1][7]


Modell von Lull
Die Bedeutung des Fossils wurde auf einem interkollegialen Treffen der geologischen Abteilungen bestätigt, und als der amerikanische Paläontologe Richard Swann Lull Talbot anschließend ermutigte, das Exemplar zu beschreiben, antwortete sie, sie wisse nichts über Dinosaurier, aber Lull schlug vor, sie zu studieren und dann beschreibe es. Im Dezember 1910 las Talbot auf dem Treffen der Paläontologischen Gesellschaft in Pittsburgh eine vorläufige Beschreibung des Fossils und veröffentlichte im Juni 1911 eine kurze wissenschaftliche Beschreibung, in der sie das Exemplar zum Holotyp der neuen Gattung und Art machte Podokesaurus Holyokensis.[1][8] Der Gattungsname leitet sich von den altgriechischen Wörtern ab podōkēs ((ποδώκης), was “schnell (oder flott)) bedeutet, ein Beiname, das üblicherweise in Bezug auf den griechischen Helden Achilles verwendet wird, und Saura ((σαύρα) bedeutet “Eidechse”, während sich der spezifische Name auf Holyoke bezieht. In vollem Umfang kann der Name als “schnellfüßige Eidechse von Holyoke” übersetzt werden.[8][2] Die Entdeckung und Benennung von Podokesaurus machte Talbot zur ersten Frau, die einen Dinosaurier fand und beschrieb.[9] Der amerikanische Paläontologe Robert T. Bakker erklärte 2014, während alte Professoren murrten, dass Frauen während seiner Zeit an der Universität nicht für die Arbeit mit Fossilien geeignet seien, entdeckte Talbot Podokesaurus war ein Gegenargument dazu.[10]
Zum Zeitpunkt der Veröffentlichung der Beschreibung hatte Talbot das Fossil zur weiteren Vorbereitung und Untersuchung an das Peabody Museum of Natural History der Yale University geschickt, wo auch gegossene Repliken der Knochen angefertigt wurden, die im Felsen lagen. Dort zeichnete Lull eine Rekonstruktion des Skeletts, wobei die im Fossil fehlenden Teile auf den Äquivalenten in beruhten Compsognathusund schuf ein Modell des Tieres im Leben, das Talbot später als “sardonisches Lächeln” bezeichnete. Lull erweiterte Talbots Artikel in einer Veröffentlichung von 1915, in der er auch andere Identitäten für einige der Knochen vorschlug als ursprünglich vorgeschlagen, einschließlich eines partiellen Coracoid anstelle eines Schulterblatts und eines Teils der Tibia anstelle des Ilium (Teil der Hüften). Lull vermutete, dass der Felsbrocken, der das Fossil enthielt, von einem Felsvorsprung stammen musste, der sich auf der Südseite der Holyoke Range befand, etwa zwei oder drei Meilen nördlich von der Stelle, an der er gefunden wurde, und gab an, dass er aus dem Longmeadow-Sandstein stammte.[6][2][11][7]

Vorderer Teil des Holotyps

Interpretationszeichnung von 1911
Lull hatte sein Manuskript vor der Veröffentlichung zur Kritik an den dänischen Ornithologen Gerhard Heilmann geschickt. Heilmann veröffentlichte seine Antwort in einem Artikel von 1913, in dem er zuvor unveröffentlichte Fotos des von Talbot erhaltenen Fossils sowie seine eigenen Restaurierungen enthielt. Er war mit einigen anatomischen Interpretationen von Lull nicht einverstanden und hatte in Bezug auf das Fossil mit dem amerikanischen Zoologen Robert Wilson Shufeldt über seine Behauptungen korrespondiert. Heilmanns Artikel war einer aus einer Reihe über die Herkunft von Vögeln, in der er die Skelette prähistorischer Reptilien untersuchte, um Merkmale zu finden, die möglicherweise von Vögeln abstammen.[12] In einem Treffen der Washington Academy of Sciences von 1916 berichtete Shufeldt über seine Korrespondenz mit Lull, Talbot und Heilmann und stimmte letzterem in einigen seiner Kritikpunkte an Lulls Wiederherstellung von zu Podokesaurus.[13]
Talbot wollte, dass das Fossil in einer ständigen Ausstellung in Yale oder Washington bleibt, wo es “mit seiner Art” sein könnte, aber es wurde als lokales Exemplar am Mount Holyoke im alten Wissenschaftsgebäude Williston Hall aufbewahrt, wo es zu einer “Neugierde für Haustiere” wurde ” für die Schüler. Während der Weihnachtsferien von 1917 brannte Williston Hall nieder und es gab keine Überreste der Podokesaurus Fossilien wurden in den Trümmern gefunden. Der amerikanische Schriftsteller Christopher Benfey wies 2002 darauf hin Podokesaurus hatte daher den besonderen Unterschied, der Dinosaurier zu sein, der zweimal verschwand.[2][1] Der deutsche Paläontologe Friedrich von Huene war der letzte, der das Holotyp-Exemplar in einer Veröffentlichung von 1914 untersuchte und beschrieb, bevor es zerstört wurde (er hatte es in Yale untersucht, bevor Talbots Beschreibung veröffentlicht wurde).[7][14] Während die Fossiliensammlungen des Colleges durch das Feuer fast vollständig zerstört wurden, wuchsen seine Einrichtungen und Sammlungen aufgrund von Talbots Bemühungen weiter und verbesserten sich danach.[15]
Keine andere eindeutige Podokesaurus Seitdem wurden Exemplare gefunden, aber gegossene Nachbildungen des Typusexemplars verbleiben im Peabody Museum of Natural History und im American Museum of Natural History in New York. Podokesaurus erhielt wenig weitere Aufmerksamkeit, bis neue und reichlich vorhandene Fossilien von Coelophysis wurden Ende der 1940er Jahre entdeckt und die Anatomie kleiner Theropodendinosaurier wurde klarer; Die amerikanischen Paläontologen Edwin Harris Colbert und Donald Baird verglichen Podokesaurus mit Coelophysis im Jahr 1958 unter Verwendung der Abgüsse der ersteren. Zusammen mit dem Sauropodomorph Anchisaurus, Podokesaurus war eine der ersten bedeutenden Sammlungen von Dinosaurierskelettmaterial, die an der Ostküste Nordamerikas bekannt waren. Abgesehen von einem Zahn (zugeordnet zu Coelophysis 1976) sind alle anderen Theropoden-Aufzeichnungen des frühen Jura an der Ostküste Fußabdrücke.[16][17][18]
Beschreibung[edit]
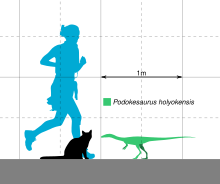
Der erhaltene Körper des einzig bekannten Podokesaurus Das Fossil war 18 cm lang.[8] Lull schätzte die gesamte Länge von Podokesaurus 1.150 mm (3,77 ft) gewesen sein, der Hals 130 mm (5,1 in), der Rücken 170 mm (6,7 in), der Kreuzbein (die Wirbel der Beckenregion) für 45 mm (1,8 in) und der Schwanz für 730 mm (2,40 ft).[6] Von Huene schätzte das Tier auf eine Länge von 1.100 mm (3,6 ft), wobei der Schwanz etwa 70 cm (2,3 ft) ausmachte, mehr als das 1,5-fache des restlichen Skeletts zusammen.[7] Colbert schätzte die Länge des Tieres auf etwa 1 m.[17] 1995 schätzte der Schriftsteller Jan Peczkis Podokesaurus 10–40 kg (22–88 lb) durch Bestimmung der Beckenhöhe (basierend auf von Huenes Messungen) gewogen zu haben.[19] Der Paläontologe Gregory S. Paul schätzte das Gewicht 2016 auf 1 kg (basierend auf einer Schätzung von 1 m Länge).[20] Talbot gab 1937 an, dass die Wissenschaftler, die das Exemplar gesehen hatten, nicht glaubten, es sei ein junges Individuum, da es keine Hinweise auf Knorpel gab, die sich mit zunehmendem Alter in Knochen verwandeln würden.[1] während Paul dachte, es sei möglicherweise jugendlich.[20] Im Allgemeinen war das Tier leicht konstruiert und hatte hohle Knochen.[8] Als Coelophysoid wäre es wahrscheinlich ähnlich aufgebaut wie zum Beispiel Coelophysis, schlank, langhalsig und mit scharfen, zurückgebogenen Zähnen.[16]
Da waren damals nur wenige andere kleine Theropoden bekannt Podokesaurus beschrieben wurde, wurde es hauptsächlich mit den späteren Gattungen verglichen Compsognathus, Ornitholestes, und Ornithomimus, die für verwandt gehalten wurden. Aufgrund der fragmentarischen Natur der Podokesaurus Probe und die Tatsache, dass es verloren geht, die Identität und Form einiger seiner Knochen sind heute unklar. Da nur noch Abgüsse des Exemplars übrig sind, kann nur ein allgemeiner Eindruck vom erhaltenen Skelett und seinen Proportionen gewonnen werden. Talbot identifizierte Knochenfragmente neben dem Schwanz als Schädelknochen, da zwei von ihnen bilateral symmetrisch waren und eines breit konvex mit einem Sulkus (oder einer Furche) in der Mittellinie war und laut Lull möglicherweise an der oberen Schädelwand platziert wurde , nicht weit von der Stirnknochen. Lull dachte, andere dieser Elemente könnten Teil des gewesen sein Nasenknochen.[6][17] Von Huene interpretierte diese Knochen stattdessen als Schwanz aus der Mitte des Schwanzes.[7]
Postkranielles Skelett[edit]

Interpretationszeichnung des Teilschwanzes und unsicherer Knochenfragmente (oben links)
Querschnitt eines Wirbels
Die Wirbel waren sehr leicht und hohl und einige waren an jedem Ende leicht konkav. Die Halswirbel waren im Vergleich zu den Rückenwirbeln relativ groß in Länge und Durchmesser und hatten stilförmige (einem Stift oder Borsten ähnelnde) Halsrippen, die lang, gerade und schmal waren, wie in Anchisaurus. Die vorderen Rücken waren viel kürzer als die Halswirbelsäule und nahmen dann zum Becken hin wieder zu. Sie hatten eine Länge von 12–13 mm (0,47–0,51 in). Das Nervenstacheln der Rücken waren schmal, hoch und nach hinten gebogen, aber nicht so von vorne nach hinten erweitert wie in Compsognathus. Die Rückenrippen waren stark gebogen und sehr schlank und hohl, was an die in erinnerte Saltopus und HallopusDas größte ist 52 mm lang und 2 mm breit. Das Querprozesse (die mit den Rippen auf jeder Seite eines Wirbels verbunden waren) bildeten einen Kreisbogen von jeder Seite eines Rückens nach unten, der sich ungefähr so weit von der Seite des Wirbels erstreckte Zentrum (der “Körper” eines Wirbels) als der Dornfortsatz war hoch. Das Zygapophysen (Prozesse, die benachbarte Wirbel verbanden) waren gut entwickelt. Die Zentra waren relativ länger als die von Ornithomimus, während die von Compsognathus ähnelte denen von Podokesaurus mehr, waren aber kürzer.[8][6][7]
Während sich der hintere Teil des Schwanzes in einiger Entfernung vom Rest des Skeletts befand, glaubte Lull, dass er sich in seiner natürlichen Position befand und dass er sehr lang gewesen wäre, da die Zwischenwirbel weggefegt worden waren. Die Schwanzwirbel waren sehr lang und schlank und zusammen mit ihren Chevrons (Knochen an den Unterseiten der Caudalen) ähnelten denen von Ornitholestes. Ein typischer Schwanz war 17 mm lang und 4 mm im Durchmesser. Die Caudals waren sehr ähnlich lang und nahmen erst in den letzten paar aufeinanderfolgenden Längen ab. Direkt vor dem Schambein befand sich eine Ansammlung von Schlanken Gastralia (Bauchrippen), von denen die längste 18 mm lang war. Die vorderste Gastralia hatte verbreiterte Enden, was häufig bei sternalen Rippen der Fall ist.[8][6][7]

Der Humerus war klein und sehr empfindlich, 42 mm lang und weniger als halb so lang wie der Femur. Es hatte einen starken Processus lateralis (ein zur Seite gerichteter Prozess) 30 mm über seinem unteren Ende. Lull fand es ähnlich wie das von Ornitholestes, abgesehen von dem radialen Kamm (der mit dem verbunden ist Radius Knochen des Unterarms) nicht so hoch ist und wenig Muskelkraft zeigt. Von Huene schätzte den Radius auf 30 mm. Lull fand die Hand der von ähnlicher Compsognathus als von Ornitholestes.[8][6][7] Es blieben nur schlanke Phalanxknochen der drei Finger erhalten, einschließlich scharfer, gebogener Krallen.[7] Das Schambein war sehr lang und dehnte sich sowohl am vorderen als auch am hinteren Ende aus. Es war 6 mm (0,24 Zoll) breit und 95 mm (3,7 Zoll) lang, und seine geringere Ausdehnung hatte einen Durchmesser von 10 mm (0,39 Zoll). Talbot fand die Scham ähnlich der der damals unbeschriebenen Procompsognathus. Lull argumentierte, dass die Schambein ungefähr im Fossil lag und nach vorne zeigte.[8][6] Heilmann glaubte stattdessen, der Knochen sei vor der Beerdigung aus seiner Position gebracht worden und wäre nach hinten gerichtet gewesen.[12] Colbert und Baird schlugen vor, dass das Schambein nach vorne gerichtet war, sich jedoch aufgrund natürlicher Verwerfungen und Knochenrisse leicht nach oben und nicht nach unten krümmte. Das Ischium war 55 mm lang, am dem Körper am nächsten gelegenen Ende 15,5 mm breit, und sein schlanker Schaft hatte einen Durchmesser von 4 mm und war am unteren Ende etwas verdickt. Lull fand es ähnlich wie bei Compsognathus und Ornitholestes. Unter dem Acetabulum (wo der Femur mit dem Becken verbunden ist) gab es eine breite Lamelle, die zum Schambein lief.[6][7]
Der Femur war schlank, fast gerade, hatte dünne Wände und war an der Rückseite seines unteren Endes erweitert. Es war 86 mm lang und hatte einen Durchmesser von 6,5 mm knapp unterhalb des vierter Trochanter (Ein Flansch in der Mitte der Länge auf der Rückseite des Femurs). Der vierte Trochanter war stark, 18 mm lang und etwa 2 mm hoch und befand sich hinter der Mitte des Schafts zum unteren Ende hin. Die Kondylen (runde Vorsprünge) des unteren Endes des Femurs ragten stark nach hinten heraus. Die Tibia war ein dünner, schmaler Schaft mit einer Länge von 104 mm (4,1 Zoll) und einem Durchmesser von etwa 7 mm (0,28 Zoll). Talbot sagte das sehr dünn Fibula lag nahe an der Tibia und war fast gleich lang, während von Huene dachte, dies sei stattdessen Teil der zerquetschten Tibia. Die drei Mittelfußknochen des Unterschenkels waren eng zusammengedrückt und bildeten eine kompakte Struktur, ähnlich wie in Ornithomimus, aber nicht zu einem Tarsometatarsus verschmolzen, wie bei Vögeln zu sehen. Die Länge der Mittelfußknochen betrug 75 mm (3,0 Zoll), während ihre Breite 9 mm (0,35 Zoll) betrug. Die vollständigste Pedal-Phalanx war sehr schlank und 12 mm lang.[6][8][7]
Einstufung[edit]
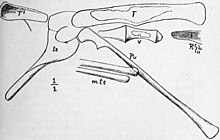
Als Talbot 1910 ihre vorläufige Beschreibung las, schlug er vor Podokesaurus ein “pflanzenfressender Dinosaurier” zu sein, aber weitere Arbeiten an der Yale University zeigten, dass einige Knochen falsch identifiziert worden waren und das Ischium des Beckens mit einem gut entwickelten Kamm dem von ähnelte Compsognathus. Sie verzichtete auf eine eindeutige Klassifizierung des Exemplars in ihrer Beschreibung von 1911, da Kiefer- und Fußknochen fehlten, die dies hätten unterstützen können, kam jedoch zu dem Schluss, dass es aufgrund seiner Form und Position zu einem “fleischfressenden Dinosaurier” gehört hätte das Schambein sowie das Fehlen eines Postpubis. Sie betrachtete das Fossil als triasisch (die erste der drei mesozoischen Perioden).[8] 1914 nannte von Huene die neue Familie Podokesauridae, in der er zusätzlich zu Podokesaurus, inbegriffen Saltopus, Procompsognathus, Coelophysis, und Tanystropheus. Er stellte diese Gruppe unter Coelurosauria, zu der zu dieser Zeit im Allgemeinen kleine Theropoden gehörten.[21][7][22]

Heilmann überlegte Podokesaurus sehr ähnlich dem frühen Vogel Archaeopteryx im Jahr 1913 und fragte sich, warum Talbot keine Vergleiche damit gemacht hatte. Er fand insbesondere die Beine und das Becken, als der Schambein so interpretiert wurde, dass er nach hinten zeigte (wie bei Vögeln), ähnlich wie es bei einem Vogelvorfahren aussehen könnte, während er Lulls Rekonstruktion eines nach vorne gerichteten Schambeins für unwahrscheinlich hielt. Er fand auch Teile des Schulterblatts, der Vorderbeine und sogar der Schwanzknochen, die denen von Vögeln ähnlich waren. Er wies außerdem auf das Vorhandensein von Bauchrippen hin und darauf, dass die Knochen hohl seien, und kam zu dem Schluss, dass nichts ausgeschlossen sei Podokesaurus von einem sehr primitiven Protovogel mit noch nicht verschmolzenen Mittelfußknochen. Das einzige Merkmal, das er als unvereinbar mit dieser Interpretation empfand, war der kurze Finger mit einer Kralle, wenn er nicht stattdessen Teil eines Zehs war. Er schlug vor, dass das Problem festgestellt werden könnte, wenn der Schädel, das Brustbein und Schlüsselbein wurden im Felsen Knochen gefunden, die wichtige Hinweise für die Klassifizierung sein würden. Heilmann wünschte sich eine klarere Erklärung für die Platzierung der Mittelfußknochen im Verhältnis zueinander und fand es überraschend, dass Lulls Rekonstruktion von Podokesaurus zeigte einen noch stärker angedrückten Mittelfuß als der später vermutete Nachkomme Ornithomimus, von dem er dachte, dass er zu diesem Zeitpunkt einen völlig vogelartigen Mittelfuß entwickelt hätte. Heilmann gefunden Podokesaurus Sehr ungewöhnlich für einen Dinosaurier aus der Trias und dachte, es sei eines der frühesten zweibeinigen Wirbeltiere. Er schlug das vor, da es so anders war als Thecodontosaurus und Plateosaurus, das können keine Dinosaurier gewesen sein, oder Podokesaurus könnte stattdessen verwandt sein mit Saltopus oder Parasuchianer, fand aber seine Fossilien zu unzureichend, um etwas Bestimmtes über das Thema zu sagen.[12]
Shufeldt ging 1915 auf seine und Heilmanns Interpretation des Schambeins ein und stellte fest, dass der Knochen wahrscheinlich während der Fossilisierung verschoben worden war. Er wies darauf hin, dass es, wenn es nach vorne gerichtet gewesen wäre, in gewaltsamem Kontakt mit den Bauchrippen gewesen wäre, was für die inneren Organe während der Bewegung gefährlich gewesen wäre.[11] Lull fand Heilmanns und Shufeldts Kritik an seiner Beckenrekonstruktion wahrscheinlich im Jahr 1915. Er listete Podokesauridae unter der Überfamilie Compsognatha auf und schlug vor Podokesaurus war vielleicht Vorfahr der späteren nordamerikanischen Gattungen Ornitholestes und Ornithimimus.[6] Im Jahr 1916 wies Heilmann darauf hin, dass frühe Dinosaurier, Parasuchier und Flugsaurier alle Ähnlichkeiten mit Vögeln und untereinander hatten und dass Trias-Reptilien dies mögen Scleromochlus, Saltopus, und Podokesauruswaren schwer zu trennen. Er schlug das vor Podokesaurus und andere zweibeinige Reptilien aus der Trias stammten von Pseudosuchianern ab (von denen viele Arten von Archosauriern zu dieser Zeit klassifiziert wurden), die möglicherweise eine Bipedalität durch das Perm entwickelt haben. Er kam zu dem Schluss, dass Vögel aufgrund ihrer Zweibeinigkeit und vogelähnlichen Beine eher von ornithosuchischen Pseudosuchianern als von Dinosauriern abstammen.[23] Der Paläontologe Henry Fairfield Osborn verwendet Podokesaurus als Beispiel für einen “fleischfressenden Dinosaurier vom vogelartigen Typ” im Jahr 1917, wobei festgestellt wurde, dass Ähnlichkeiten zwischen Vögeln und Dinosauriern auf eine parallele Entwicklung zurückzuführen sind.[24] Der Paläontologe Franz Nopcsa von Felső-Szilvás gründete 1928 die Unterfamilie Podokesaurinae, zu der er gehörte Podokesaurus, Procerosaurus, Saltopus, und Tanystropheus.[25] Von Huene gruppiert Podokesaurus, Coelophysis, Spinosuchus, Halticosaurus, Saltopus, Avipes, und Velocipes in Podokesauridae im Jahr 1932.[26]

1958 beschrieben Colbert und Baird ein Theropoden-Exemplar, das aus natürlichen Knochenabgüssen in Sandstein bestand (wahrscheinlich gebildet, als die Knochen durch saures Wasser aufgelöst wurden und Schimmelpilze der Knochen hinterließen), einschließlich Schambein, Tibia und einiger Rippen. Sie fanden das Exemplar ähnlich wie Coelophysis und Podokesaurusund bezeichnet es als C. sp. (unbekannter Arten). Sie betrachteten die natürliche Besetzung und Podokesaurus aus der Portland Arkose der Newark Group im Connecticut Valley zu sein. Obwohl Podokesaurus war kleiner als die anderen (vergleichbar mit der kleinsten bekannten Coelophysis Proben) schlugen diese Forscher vor, dass dies, weil es ihnen so ähnlich war, Fragen nach seiner Gültigkeit aufwirft.[27][28] Im Jahr 1964 synomymisierte Colbert Podokesaurus mit Coelophysis, (da der letztere Name älter war), prägte die neue Kombination C. Holyokensis. Er schlug auch vor, dass die natürliche Besetzung dazu gehörte C. Holyokensis. Colbert erklärte das Podokesaurus konnte nur unterschieden werden von Coelophysis durch die Nervenstacheln seiner Wirbel, die von vorne nach hinten nicht so lang sind, und weil sich das Ischium in der Form unterschied. Er fand, dass diese Unterschiede denen ähneln, die zwischen modernen Reptilienarten derselben Gattung beobachtet wurden, und dass sie östliche (C. Holyokensis) und westlich (C. bauri) Arten der gleichen Gattung. Während er zugab, dass diese Schlussfolgerungen nicht ironisch waren, und das Podokesaurus Möglicherweise war dies eindeutig, sagte er, die Beweislast sollte bei den Befürwortern einer solchen Ansicht liegen.[17]
1977 haben die Paläontologen Paul E. Olsen und Peter Galton die Newark Supergroup (zu der die Portland-Formation gehört) anstelle der vorhergehenden Trias, wie zuvor angenommen, auf das frühe Jura umgestellt. Dies war Teil einer Studie, in der Olsen die Fauna verschiedener Formationen verglich und zu dem Schluss kam, dass es an der Grenze zwischen Trias und Jura kein plötzliches, weit verbreitetes Aussterben gegeben hatte, sondern dass es stattdessen allmählich gewesen war.[29] Olsen erklärte 1980, dass während die genaue Herkunft und systematische Position von Podokesaurus wird wahrscheinlich ungewiss bleiben, die Portland-Formation, von der angenommen wurde, dass sie stammt, war wahrscheinlich im frühen Jura-Alter und daher 15 Millionen Jahre jünger als Coelophysis aus der Trias. Da Funktionen zwischen geteilt Podokesaurus und Coelophysis sind auch in anderen kleinen Theropoden bekannt, wie z SyntarsusOlsen schlug das vor Podokesaurus sollte als separate Gattung unbestimmter Theropoden beibehalten werden, um eine Überdehnung des zeitlichen Bereichs von zu vermeiden Coelophysis. Er wandte dieses Argument auch auf die natürliche Besetzung an und betrachtete es als incertae sedis (mit unsicheren Beziehungen) zwischen Theropoden und nicht als Podokesaurus oder Coelophysis.[30]
Der Paläontologe Samuel P. Welles erklärte 1984, dass die Familie Podokesauridae für die meisten Trias-Theropoden zu einem “Allheilmittel” geworden sei. Während er fand Coelophysis am ähnlichsten zu Podokesaurus Bei den Theropoden waren die beiden seiner Meinung nach sehr unterschiedlich, da der vierte Trochanter des letzteren unterhalb der Mittelhöhe des Femurs lag (im Gegensatz zur höheren Position bei den meisten anderen Theropoden) und seine Mittelfußknochen gleich lang waren.[14] Der Paläontologe Kevin Padian erklärte 1986, dass Colberts Vorschlag der Synonymie zwar möglich sei, die erkennbaren Ähnlichkeiten zwischen Podokesaurus und Coelophysis waren primitive Theropodenmerkmale, und die beiden waren zeitlich nicht so nah wie gedacht.[31] Paul sagte 1988, dass während der Familie Podokesauridae noch für verwendet wurde Coelophysis und Verwandte, Podokesaurus Zunächst beruhte er nicht auf guten Überresten, und da nur noch schlechte Nachbildungen übrig waren, dachte er, dass es immer Meinungsverschiedenheiten über das Taxon geben würde. Er fand dies zu vieldeutig, um es zu ertragen, und schlug vor, stattdessen den Namen Coelophysidae zu verwenden (Halticosauridae, ein weiterer Anwärter, basierte ebenfalls auf zu fragmentarischen Überresten).[32] Im Jahr 1989 bezweifelte Colbert auch die Synonymie von Podokesaurus mit Coelophysisund dass die natürlichen Abgüsse aufgrund ihres überarbeiteten frühen Jurazeitalters zu letzteren gehörten und er keinen Versuch unternahm, den Status von zu klären Podokesaurus aufgrund des einzigen verlorenen Exemplars. Er behielt den Namen Podokesauridae für die Familie.[33]

1990 betrachteten die Paläontologen Timothy Rowe und Jacques Gauthier Podokesauridae als taxonomischen Abfallkorb, in dem Taxa nach phänetischer Ähnlichkeit und stratigraphischer Teilung gruppiert worden waren und daher ständig überarbeitet und instabil waren. Sie hielten es für möglich, dass Podokesaurus und die natürlichen gegossenen Exemplare waren Coelophysis, fanden aber heraus, dass ihre Ähnlichkeiten nicht ausschließlich von ihnen geteilt wurden, sondern Ahnenmerkmale unter den Theropoden waren. Sie waren sich daher einig, dass der Name Podokesaurus sollte auf den Holotyp beschränkt sein, und dass es und die natürliche Besetzung als Theropoda betrachtet werden sollten incertae sedis.[34] Im gleichen Band stimmte der Paläontologe David B. Norman dieser Einschätzung zu und gab die Merkmale an, die zur Vereinigung verwendet wurden Podokesaurus mit Coelophysis Bestätigen Sie lediglich, dass sie Dinosaurier waren, anstatt eine spezifische Beziehung zwischen ihnen herzustellen.[5] Der Paläontologe Thomas R. Holtz stimmte 1994 zu, dass der Familienname Podokesauridae durch Coelophysidae ersetzt werden sollte. Zu diesem Zeitpunkt geriet die Idee, kleine Theropoden in Coelurosauria und große Theropoden in Carnosauria zu gruppieren, in Ungnade, und Coelophysoidea wurde als separate Gruppe graziler, früher Theropoden angesehen.[35] Die Paläontologen David B. Weishampel und Luther Young schlugen ceratosaurische Affinitäten für vor Podokesaurus in 1996.[36]
Der Paläontologe Ronald S. Tykoski und Rowe stellten 2004 fest, dass während Podokesaurus hatte coelophysoide Merkmale (wie eine kleine, knaufartige Ausdehnung im unteren Teil des Schambeins), es hatte keine abgeleiteten Merkmale, die es vereinen würden Coelophysis. Sie stimmten früheren Forschern zu, dass der Name Podokesaurus sollte auf den Holotyp beschränkt sein und zu dem Schluss kommen, dass der natürliche Abguss ein Coelophysoid sein kann, aber nicht mehr als ein Theropod identifiziert werden kann.[37] Im Jahr 2004 gaben die Paläontologen Matthew T. Carrano und Scott D. Sampson dies an Podokesaurus war mit ziemlicher Sicherheit ein Coelophysoid ähnlich Coelophysis. Es hatte coelophysoide Merkmale wie einen langen, nach unten gekrümmten Schambein, der länger als das Ischium war, und ein zusätzliches Foramen (Öffnung) am Schambein unterhalb des Schambeins Oburator Foramen, aber über seine Affinitäten konnte wenig mehr gesagt werden.[38] Carrano und Kollegen gaben 2004 an, dass Segisaurus und Podokesaurus gehörten zu den jüngsten überlebenden Coelophysoiden, und dass die evolutionäre Strahlung dieser Gruppe möglicherweise durch den neuesten Teil des frühen Jura beendet wurde.[39] Ebenfalls im Jahr 2004 wurden die Paläontologen Anthea Bristowe und Michael A. Raath aufgeführt Podokesaurus als Synonym für Coelophysis ohne Ausarbeitung.[40] Tykoski lehnte in seiner Dissertation von 2005 die Synonymie zwischen den beiden ab, ging aber Podokesaurus aus seiner Analyse.[41] Paul schlug dies 2016 vor Podokesaurus kann die Familie Coelophysidae mit gebildet haben Coelophysis, Panguraptor, und Procompsognathus.[20]
Paläobiologie[edit]

Talbot schlug vor, dass der kurze, schlanke Humerus, die langen, geraden Hinterbeinknochen und der gut entwickelte vierte Trochanter des Femurs darauf hinwiesen Podokesaurus war zweibeinig. Sie fand die Tatsache, dass die Tibia viel länger als der Femur war, dass die Mittelfußknochen sehr lang waren, über die halbe Länge der Tibia und die leichte Konstruktion des Skeletts auf eine schnelle Fortbewegung hinwies. Talbot erklärte, dass Lull dies für eine Anpassung an die klimatischen Bedingungen hielt, da das Tier in seiner semi-ariden Region schnell und weit nach Wasser reisen musste. Talbot berichtete auch über ein kleines Stück glatten, polierten Quarzes zwischen den Rippen und vermutete, dass es sich um einen Gastrolithen (Magenstein) handeln könnte. Daher wurden diese erstmals in Verbindung mit einem fleischfressenden Dinosaurier gefunden.[8] Von Huene stimmte zu, dass der Stein ein Gastrolith war, der sich vom umgebenden Sandstein unterschied, und fügte hinzu, dass er 11 mm lang war.[7] Lull stellte fest, dass das Tier im Wesentlichen ein schlankes, flüchtiges (für das Laufen angepasstes) Tier mit fleischfressenden Gewohnheiten war, aber dass die Schlankheit, die es schnell machte, es auch auf kleine Beute beschränkte.[6] 1932 schlug von Huene vor, dass kleine Coelurosaurier einen springenden Gang haben, da ihre Unterschenkel länger als ihre Oberschenkel sind, im Gegensatz zu den abwechselnden Schritten der Carnosaurier. Mit seinem großen vierten Trocanther, dachte er Podokesaurus hatte wahrscheinlich diesen springenden Gang aufgegeben und sich stattdessen mit schnellen, abwechselnden Schritten ähnlich wie Laufvögel bewegt.[26]
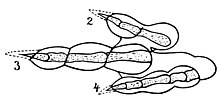
1982 schätzte der Paläontologe Richard A. Thulborn die Geschwindigkeit verschiedener Dinosaurier auf der Grundlage der Beziehungen zwischen Geschwindigkeit, Gang und Körpergröße moderner Tiere (hauptsächlich Säugetiere). Durch Extrapolation der Schrittlänge und Trittfrequenz von PodokesaurusEr schätzte, dass es mit etwa 15 bis 20 km / h gelaufen sein könnte. Er kam zu dem Schluss, dass ein Dinosaurier, wenn er in der Geschwindigkeit mit einem Säugetier mithalten kann, eine ähnliche Effizienz in Bezug auf Bewegungsfähigkeit und Physiologie aufweisen muss. Er warnte davor, dass der Vergleich schwierig sei, da sich die Anatomie der Dinosaurier erheblich von der von Säugetieren und Vögeln unterscheide. Zu den Unterschieden gehörten die massiven Schwänze von Dinosauriern und die Retraktormuskeln der Hinterbeine, die hinter dem Femur entstanden, während diese Muskeln bei Säugetieren und vorhandenen Vögeln weiter nach vorne gerichtet waren.[42] Weishampel erklärte im Jahr 2006, dass als Coelophysoid, Podokesaurus wäre ein Raubtier mit leichtem Fuß gewesen, mit mächtigen Vorderbeinen und greifenden Händen.[16]
Lull fand, dass fossile Fußabdrücke benannt wurden Grallator cursorius stimmte mit dem Fuß und Schritt von Podokesaurus. Diese waren in einem Steinbruch in South Hadley reichlich vorhanden (wo sie neben großen gefunden wurden Otozoum Moodii Tracks), von denen er glaubte, dass sie diese Identität unterstützen.[6] Im Jahr 1926 schlug Heilmann vor, dass der Fuß von Procompsognathus war ein besseres Spiel für die G. cursorius Spuren, aufgrund der Proportionen seiner Zehenknochen.[43] Der Paläontologe Wilhelm Bock erklärte 1952, dass dies zwar sehr ähnlich sei Grallator Spuren waren verschiedenen kleinen Dinosauriern zugeschrieben worden (wie z Podokesaurus) hielt er solche Korrelationen für zu spezifisch, und das Beste, was gesagt werden konnte, waren solche Spuren, die kleine Coelurosaurier darstellten.[44] Der Schriftsteller Donald F. Glut schlug 1997 vor, eine Platte mit Spuren aus der Brunswick Formation von New York zuvor zuzuschreiben Coelophysis kann stattdessen von produziert worden sein Podokesaurus, basierend auf seinem Jurazeitalter.[45] Diese Formation ist heute als Passaic-Formation bekannt und wird stattdessen als spättriassisch angesehen.[46]
Paläoumgebung[edit]
Das einzige bekannte Exemplar von Podokesaurus Es wird angenommen, dass es aus der Portland-Formation (früher bekannt als Portland Arkose) im Hartford Basin von Massachusetts stammt. Das Alter dieser Formation war lange Zeit unklar (es wurde angenommen, dass es bis 1977 triassisch war), aber es wird derzeit angenommen, dass es auf die hettangisch-sinemurischen Stadien des frühen Jura vor 201 bis 190 Millionen Jahren zurückgeht (früher angenommen) die Pliensbachian-Toarcian Stadien).[47][16][30] 2016 schlugen der Paläontologe Robert E. Weems und Kollegen vor, die Portland-Formation zu einer geologischen Gruppe innerhalb der Newark Supergroup (als Portland-Gruppe) zu erheben und damit den früheren Namen “Agawam Group” zu ersetzen. Sie stellten auch den Longmeadow Sandstone wieder her, wo Podokesaurus wurde als Formation gefunden (innerhalb der obersten Portland-Gruppe); es war früher als identisch mit der Portland-Formation angesehen worden.[47][6]
Die Portland Group repräsentiert den obersten Teil der Newark Supergroup und wurde hinterlegt, nachdem die magmatische Provinz des Zentralatlantiks am Ende der Trias und zu Beginn des Jura gebildet worden war. Der Langwiesen-Sandstein besteht aus fluvialen (durch Flüsse und Bäche abgelagerten) roten Schlammsteinen, Sandsteinen, Konglomeraten, kleinen roten Äolischen (durch Wind abgelagerten) Sandsteinen und Schlicksteinen.[47] Andere aus der Formation bekannte Tiere sind der Sauropodomorph-Dinosaurier Anchisaurus, der Crocodylomorph Stegomosuchusund Fische wie Acentrophorus und Semionotus.[30] Zu den Dinosaurierspuren gehört die Ichnogenera Anchisauripus, Anomoepus, Eubrontes, und Grallator.[48]
Siehe auch[edit]
Verweise[edit]
- ^ ein b c d e f Warner, FL (1937). “XII. Verlorener Dinosaurier”. Auf einem New England Campus. Cambridge: Houghton Mifflin Company. p. 279. ASIN B00085TO0O.
- ^ ein b c d Benfey, C. (2002). “Vorwort:” Ein Weg der Vergänglichkeit“”“”. Perspektiven ändern: Der Blick vom Mount Holyoke. Ithaka: Cornell University Press. p. 9. ISBN 978-0801441196.
- ^ “Mut zu graben: Mignon Talbot (1869-1950)”. museumoftheearth.org. Abgerufen 14. August 2020.
- ^ Rossiter, MW (1981). “Geologie in der Frauenbildung des 19. Jahrhunderts in den Vereinigten Staaten”. Zeitschrift für geologische Bildung. 29 (5): 228–232. Bibcode:1981JGeoE..29..228R. doi:10.5408 / 0022-1368-29.5.228.
- ^ ein b Norman, DB (1990). “Problematische Theropoda”. In Weishampel, DB; Osmolska, H.; Dodson, P. (Hrsg.). Die Dinosaurier (1. Aufl.). Berkeley: University of California Press. p. 298. ISBN 978-0-520-06727-1.
- ^ ein b c d e f G h ich j k l m n Lull, RS (1915). “Trias Leben im Connecticut Valley”. Staatliche geologische und naturhistorische Untersuchung von Connecticut. 24: 155–169. doi:10.5962 / bhl.title.70405. ISBN 978-1167291937.
- ^ ein b c d e f G h ich j k l m von Huene, F. (1914). “Beiträge zur Geschichte der Archosaurier”. Geologische und Paläontologische Abhandlungen (auf Deutsch). 13: 1–53.
- ^ ein b c d e f G h ich j k Talbot, M. (1911). “”Podokesaurus Holyokensis, ein neuer Dinosaurier aus der Trias des Connecticut Valley “. American Journal of Science. s4-31 (186): 469–479. Bibcode:1911AmJS … 31..469T. doi:10.2475 / ajs.s4-31.186.469.
- ^ Turner, S.; Burek, Lebenslauf; Moody, RTJ (2010). “Vergessene Frauen in einer ausgestorbenen saurischen (Männer-) Welt”. Geological Society, London, Sonderpublikationen. 343 (1): 111–153. Bibcode:2010GSLSP.343..111T. doi:10.1144 / SP343.7. S2CID 130338204.
- ^ Bakker, RT (2014). “Eine Geschichte von zwei Compys: Was Jurassic Park habe richtig – und falsch – über die Dino-Anatomie “. Das Houston Museum of Natural Science. Abgerufen 18. August 2020.
- ^ ein b Moodie, RL (1913). “Einige jüngste Fortschritte in der Paläontologie von Wirbeltieren. II”. Der amerikanische Naturforscher. 47 (556): 248–256. doi:10.1086 / 279347. ISSN 0003-0147. JSTOR 2455799.
- ^ ein b c Heilmann, G. (1913–1914). “Vor nuværende viden om fuglenes afstamning. Andet afsnit: Fugleligheder blandt fortidsøgler”. Dansk Ornithologisk Forenings Tidsskrift (auf Dänisch). 8: 56–65.
- ^ Wegemann, CH; Lyon, MW (1916). “Verfahren der Akademie und der angegliederten Gesellschaften”. Zeitschrift der Washington Academy of Sciences. 6 (9): 251–259. ISSN 0043-0439. JSTOR 24521263.
- ^ ein b Welles, SP (1984). “”Dilophosaurus wetherilli (Dinosauria, Theropoda), Osteologie und Vergleiche “. Palaeontographica Abteilung A.. 185: 172.
- ^ Elder, ES (1982). “Frauen in der frühen Geologie”. Zeitschrift für geologische Bildung. 30 (5): 287–293. Bibcode:1982JGeoE..30..287E. doi:10.5408 / 0022-1368-30.5.287.
- ^ ein b c d Weishampel, DB (2006). Ein weiterer Blick auf die Dinosaurier der Ostküste Nordamerikas. III Jornadas Internacionales sobre Paleontología de Dinosaurios y su Entorno, Salas de los Infantes, Burgos, Spanien. Colectivo Arqueológico-Paleontológico Salense Actas. S. 129–168.
- ^ ein b c d Colbert, EH (1964). “Die Trias-Dinosaurier-Gattungen Podokesaurus und Coelophysis“. American Museum Novitates (2168): 1–12. hdl:2246/3350.
- ^ Galton, PM (1976). “Prosauropod Dinosaurier (Reptilia, Saurischia) von Nordamerika”. Postilla, Peabody Museum Yale Universität (169): 6.
- ^ Peczkis, J. (1995). “Implikationen von Body-Mass-Schätzungen für Dinosaurier”. Journal of Vertebrate Paleontology. 14 (4): 530–531. doi:10.1080 / 02724634.1995.10011575. ISSN 0272-4634.
- ^ ein b c Paul, GS (2016). “Theropoden”. Der Princeton Field Guide für Dinosaurier (2. Aufl.). Princeton: Princeton University Press. S. 76, 78. ISBN 978-0691167664.
- ^ von Huene, F. (1914). “Das natürliche System der Saurischia”. Zentralblatt für Mineralogie, Geologie und Paläontologie (auf Deutsch). 5: 154–158.
- ^ Padian, K. (2012). “Das Problem der Dinosaurierherkunft: Integration von drei Ansätzen zum Aufstieg der Dinosaurier”. Erd- und Umweltwissenschaftliche Transaktionen der Royal Society of Edinburgh. 103 (3–4): 423–442. doi:10.1017 / S1755691013000431.
- ^ Heilmann, G. (1916). “Vor nuværende viden om fuglenes afstamning. Femte afsnit: Førfuglen Proavis”. Dansk Ornithologisk Forenings Tidsskrift (auf Dänisch). 11: 90, 113, 137.
- ^ Osborn, HF (1917). Der Ursprung und die Entwicklung des Lebens über die Theorie des Handelns, der Reaktion und der Wechselwirkung von Energie. New York: C. Scribners Söhne. S. 210–215. doi:10.5962 / bhl.title.7397.
- ^ Nopcsa, F. (1928). “Die Gattungen der Reptilien”. Palaeobiologica (1): 163–188.
- ^ ein b von Huene, F. (1932). “Die fossile Reptilienordnung Saurischia, ihre Entwicklung und Geschichte” (PDF). Monographie zur Geologie und Paläontologie (auf Deutsch). 4 (1): 24, 35, 332.
- ^ Colbert, EH; Baird, D. (1958). “Coelurosaurier-Knochengüsse aus der Connecticut Valley Triassic”. American Museum Novitates (1901): 1–11. hdl:2246/2486.
- ^ Getty, PR; Bush, AM (2011). “Sandpseudomorphe von Dinosaurierknochen: Implikationen für die (Nicht-) Konservierung von Tetrapoden-Skelettmaterial im Hartford Basin, USA”. Paläogeographie, Paläoklimatologie, Paläoökologie. 302 (3–4): 407. Bibcode:2011PPP … 302..407G. doi:10.1016 / j.palaeo.2011.01.029.
- ^ Olsen, PE; Galton, PM (1977). “Aussterben der Trias-Jura-Tetrapoden: Sind sie echt?” Wissenschaft. 197 (4307): 983–986. Bibcode:1977Sci … 197..983O. doi:10.1126 / science.197.4307.983. PMID 17784133. S2CID 44654197.
- ^ ein b c Olsen, PE (1980). “Ein Vergleich der Wirbeltier-Assemblagen aus den Becken von Newark und Hartford (frühes Mesozoikum, Newark Supergroup) im Osten Nordamerikas” (PDF). In Jacobs, LL (Hrsg.). Aspekte der Wirbeltiergeschichte: Essays zu Ehren von Edwin Harris Colbert. Flagstaff: Museum of Northern Arizona Press. S. 35–53. ISBN 978-0897340526.
- ^ Padian, K. (1986). “Auf dem Typ Material von Coelophysis Cope (Saurischia: Theropoda) und ein neues Exemplar aus dem versteinerten Wald von Arizona (Spättrias: Chinle Formation) “. In Padian, K. (Hrsg.). Der Beginn des Zeitalters der Dinosaurier: Faunaler Wandel über die Grenze zwischen Trias und Jura. Cambridge: Cambridge University Press. S. 57–60. ISBN 978-0521367790.
- ^ Paul, GS (1988). Raubdinosaurier der Welt. New York: Simon & Schuster. S. 258–259. ISBN 978-0-671-61946-6.
- ^ Colbert, EH (1989). “The Triassic Dinosaur Coelophysis”. Museum of Northern Arizona Bulletin. 57: 24, 28–29. ISBN 978-0897340977.
- ^ Rowe, T.; Gauthier, J. (1990). “Ceratosauria”. In Weishampel, DB; Osmolska, H.; Dodson, P. (Hrsg.). Die Dinosaurier (1. Aufl.). Berkeley: University of California Press. S. 153, 166. ISBN 978-0-520-06727-1.
- ^ Holtz, TR (1994). “Die phylogenetische Position der Tyrannosauridae: Implikationen für die Theropodensystematik”. Zeitschrift für Paläontologie. 68 (5): 1100–1117. doi:10.1017 / S0022336000026706. ISSN 0022-3360. JSTOR 1306180.
- ^ Weishampel, DB; Young, L. (1996). Dinosaurier der Ostküste. Baltimore: Johns Hopkins University Press. p. 275. ISBN 9780801852176.
- ^ Tykoski, RS; Rowe, T. (2004). “Ceratosauria”. In Weishampel, DB; Dodson, P.; Osmolska, H. (Hrsg.). Die Dinosaurier (2. Aufl.). Berkeley: University of California Press. p. 68. ISBN 978-0-520-24209-8.
- ^ Carrano, MT; Sampson, Scott D. (2004). “Eine Übersicht über Coelophysoide (Dinosauria: Theropoda) aus dem frühen Jura Europas mit Kommentaren zur späten Geschichte der Coelophysoidea”. Neues Jahrbuch für Geologie und Paläontologie – Monatshefte. 2004 (9): 537–558. doi:10.1127 / njgpm / 2004/2004/537.
- ^ Carrano, MT; Hutchinson, JR; Sampson, SD (2005). “Neue Informationen zu Segisaurus halli, ein kleiner Theropodendinosaurier aus dem frühen Jura von Arizona “. Journal of Vertebrate Paleontology. 25 (4): 835–849. doi:10.1671 / 0272-4634 (2005) 025[0835:NIOSHA]2.0.CO; 2.
- ^ Bristowe, A.; Raath, MA (2004). “Ein jugendlicher Coelophysoid-Schädel aus dem frühen Jura von Simbabwe und die Synonymie von Coelophysis und Syntarsus“”. Palaeontologia Africana. ISSN 0078-8554.
- ^ Tykoski, RS (2005). Anatomie, Ontogenese und Phylogenie von coelophysoiden Theropoden. Die Universität von Texas in Austin (These). p. 69.
- ^ Thulborn, RA (1982). “Geschwindigkeiten und Gänge von Dinosauriern”. Paläogeographie, Paläoklimatologie, Paläoökologie. 38 (3–4): 227–256. Bibcode:1982PPP …. 38..227T. doi:10.1016 / 0031-0182 (82) 90005-0.
- ^ Heilmann, G. (1926). Der Ursprung der Vögel. New York: Dover-Veröffentlichungen. p. 181. ISBN 0-486-22784-7.
- ^ Bock, W. (1952). “Trias-Reptilienspuren und Trends der Lokomotivenentwicklung: Mit Anmerkungen zur Korrelation”. Zeitschrift für Paläontologie. 26 (3): 395–433. ISSN 0022-3360. JSTOR 1299951.
- ^ Glut, DF (1997). Dinosaurier: Die Enzyklopädie. Jefferson: McFarland & Company, Inc., S. 718–719. ISBN 978-0786472222.
- ^ Metz, R. (2000). “Trias-Spurenfossilien aus Lacustrine-Küstenvorkommen der Passaic-Formation, Douglassville, Pennsylvania”. Ichnos. 7 (4): 253–266. doi:10.1080 / 10420940009380165. S2CID 128688223.
- ^ ein b c Weems, RE; Tanner, LH; Lucas, SG (2016). “Synthese und Überarbeitung der lithostratigraphischen Gruppen und Formationen im Oberperm? – Unterjura-Newark-Supergruppe im Osten Nordamerikas”. Stratigraphie. 13 (2): 111–153.
- ^ Collette, JH; Getty, PR; Hagadorn, JW (2011). “Einblicke in einen Lebensraum der frühen Jura-Dinosaurier: Ichnofazien und rätselhafte Strukturen aus der Portland-Formation, Hoover Quarry, Massachusetts, USA”. Atlantische Geologie. 47: 81–98. doi:10.4138 / atlgeol.2011.003.
Recent Comments