Enterobacterien-ウィキペディア
腸内細菌 または、2016年に設立された注文の代表者 腸内細菌 バクテリアドメイン内の大きなグループです。系統発生系によると、それらは部門のガンマプロト酸バクテリアのクラスに属します( 分割 、Prokaryotも同様です 門 記述された)プロテオバクテリアとそこで独自の順序を形成します。
2016年以前には、この細菌群で「腸内細菌」という名前が使用されていましたが、これは細菌学的コード(ICBN)の規則に従って有効ではありませんでした。さらに、順序の中には、腸内菌 – 腸内菌 – のみがありました。注文の確立は、以前に知られている分類群にいくつかの新しい家族に接続されています。 [初め] たとえば、新しいシステムは属が エシェリチア 、 モルガネラ と yersinia 3つの異なる家族に属します(セクションSystematikを参照)。
Enterobacteriaという名前は導かれます 全体 ( 古代ギリシャ語 rowられた 「ダルム」)彼らの多くは典型的な腸の居住者だからです。しかし、多くの野生でユビキタスな、非腸の細菌の種類もこの順序に属します。
外観 [ 編集 | ソーステキストを編集します ]
細胞はロッド型で、通常は1〜5 µmの長さで、直径は約0.5〜1.0 µmです。内胞子は形成されていません。ほとんどは鞭毛で積極的に移動できますが、モチルですが、積極的に動くことができないジャンルも発生します。細胞壁は、数層のムレ層と2番目のリン脂質とリポ多糖類の外膜で構成されているため、腸内菌はグラム陰性です。
代謝 [ 編集 | ソーステキストを編集します ]
あなたの代謝はオプションの嫌気性であるため、酸素の存在下で酸化を介して物質を分解し、嫌気性条件下で発酵を作動させることができます(酸素なし)。個々の属を区別するために使用される2つの重要な嫌気性代謝経路は、2.3いじめと混合酸発酵です( 混合酸発酵 )。混合酸発酵には、主に酸性酸、乳酸、沈下酸(コハク酸)などの酸がありますが、ビンジオールはありません。 2.3-ブッツィンジオール発酵の場合、末端および副産物としてのグルコースの発酵は、特に大量のアルコール2.3-ブッツィンジオールでの量です。 2.3気流の発酵のもう1つの特徴は、中間アセトインとはるかに高いガス生産です(CO 2 )。肉屋の発酵zを見つけることができます。コウモリ Enterobacter 、 クレブシエラ 、 エルウィニア と セラティア 。混合酸性度などを使用します。将軍のような エシェリチア 、 サルモネラ と プロテウス 。
個々の属を決定するために、多数の診断テストが使用されています。たとえば、Voges Proskauerテストの助けを借りて、2.3気粘液発酵の中間アセトインが実証されています。酵素β-ガラクトシダーゼの検出は、しばしば分化するために使用されます。この酵素を持つ細菌は、二糖類乳糖(乳糖)を単糖のグルコースとガラクトシスに加水分解して、それらを代謝に使用できます。
Phylogenetik [ 編集 | ソーステキストを編集します ]
系統発生方法の使用は、部族の歴史と細菌間の関係によって明らかにすることができます。 2016年に公開されたenterobacteralの順序の説明では、5つの保存された特性インデル。 保存された署名インサートと削除 、CSI;順序の代表者の典型であるが、他の細菌では発生しない系統学および分類学のセクションの詳細)。 5つのCSIは、タンパク質用の遺伝子配列で発生します l – アラビン症イソメラーゼ、伸長因子-P類似タンパク質 YEIP 、コードペプチドABCトランスポーター透過腫瘍、ピロホスファターゼ、および仮想タンパク質。 [初め]

多くの腸内菌は、人間と動物の健康な腸内植物の一部です。ただし、環境(土壌、水)のいたるところにも発生します。いくつかは人間や動物の病原体です。それらはしばしば院内病原体(「病院細菌」)として発生し、免疫系が弱い人に影響を与えます。
腸内菌のおそらく最も重要な代表はです 大腸菌 、微生物学だけでなく、遺伝学と生化学の最も重要なモデル生物の1つ。属も印象的です プロテウス 、いわゆる「群れ現象」を観察します。これらの細菌のコロニーが寒天プレートに広がると、同心円状の輪が付いた細菌の芝生を見ることができます。
系統発生系が使用する方法 [ 編集 | ソーステキストを編集します ]
「Enterobacteriales」の代表者では、特に16S RRNAの検査に基づいた近年の系統発生法の結果は、リボソームRNAからの原核生物の満足のいく結果をもたらしていません。したがって、カナダのオンタリオ州のマクマスター大学(Adeolu et al。)の目標は、グループの多くの代表者と同じように遺伝的特徴の比較に基づいた系統発生系を作成するための目標でした。検査の時(2016年)に、国立バイオテクノロジー情報センター(NCBI)のゲノムデータベースで、腸内菌(NCBI)の14,000のゲノムが既に利用可能でした。分析のために、179の代表的なゲノム配列が使用され、さまざまな分子マーカーが選択され、比較されて比較されました。これは、腸内菌の代表者間の進化的関係を表す系統樹を作成しました。 [初め]
一方では、これには1548の主要なタンパク質が使用されました。これはグループの典型であり、タンパク質をコード化したDNA配列を比較しました。同様に、53のリボソームタンパク質(Rprotein)を使用し、4つのタンパク質をコードするいくつかの遺伝子座(MLSA)のDNA配列分析も実行されました。タンパク質はそうです gyrb (girase b)、 RPOB 、 ATPD と INFB 。このように構築された3つの系統発生ツリーは、7つのcladを持つ均一な写真になります。 Enterobacter-Escherichia -the Erwinia-Pantoea -the Pectobacterium-dickeya -the Yersinia-serratia -the Hafnia-Edwardsiella -the Proteus-Xenorhabdus – クレードと ブドヴィシア -ktkladeが呼び出されます。これに基づいて、この順序は7つの家族(別の家族であるThorselliaceaeが別の科学者グループによって最初に記述された)で定義されています。この系統解析の結果は、ゲノムカバーの分析の結果によってサポートされています。この方法も実行されます シリコで (コンピューターベース)DNA DNAハイブリダイゼーションの代替品として、クラス経済クラスクラスに役立ちます。 [初め]
遺伝的新規分析により、「保存された(保存)分子特性」の発見または決定も可能になります。これは、互いに関連する生物のグループ内で互いに違いもほとんど違いもありません。原核生物の系統発生系については、「保存された特徴的なインデル」。 保存された署名インサートと削除 、CSI)。グループ固有のCSIは、他のグループではなく、このグループのすべての代表者に登場するため、特定の分類群(たとえば1つの注文)の特徴です。グループ固有のCSIの元のインデルは、おそらく最後の共同祖先で分裂する前に発生し、結果のグループの代表者(「継承」)に引き継がれました。 [2] Adeoluらによる検査のもう1つの重要な結果。 71 CSIの発見であり、5つを含むもので、注文のすべての代表者にとってユニークなものであるため、説明に使用されます。残りの66 CSIは7つのグループに固有のものであり、体系的な分類のために、新たに発見された腸内細菌の将来の遺伝小説に使用できます。 [初め]
| 注文内の関係へのクラドグラムEnterobacter(2016年現在) [初め] |
|---|
| クラドグラムは、関連する特性インデル(CSI)の数を示します。 Thorselliaceaeの家族は含まれていません。 |
2016年の広範なゲノム全体の分析の結果は、以下に示すシステムです。
命令の命名法と分類 [ 編集 | ソーステキストを編集します ]
属 Enterobacter 腸内菌のタイプではありません、これは属です エシェリチア 。これは、に従って指定された命名法を意味します 細菌学的コード (バクテリアの国際命名法)逸脱。1958年には 司法意見15 司法委員会 (ほぼ「司法または公平な委員会」) 原核生物の体系的委員会 (原核生物の系統学に関する国際委員会、ICSP)が確認されました。 [3] 細菌学的コードの規則に従って、タイプのタイプの順序 エシェリチア したがって、「Escherichiales」と呼ばれる、タイプのタイプの順序 Enterobacter 「Enterobacterales」と呼ぶ必要がありますが、「Enterobacterial」ではありません。 [4] 新しいシステムの受け入れを確保し、秩序を「大極性」と名付けたときの混乱の可能性を回避するために、Enterobactteralesという名前が叙階されました。 11月タイプのジャンルで Enterobacter 選ばれた。 [初め]
現在のシステム [ 編集 | ソーステキストを編集します ]

2016年のEnterobacteralesの確立された順序には、合計60属の8つの家族が含まれています。 [初め] [5] [6] 新しく定義されている、したがって、細菌学コード(ICBN)のルールは、順序の型に対応するジャンルです Enterobacter 。注文腸内菌の説明 ADORD et al。 2016 ord。 11月2005年のベルゲイの系統的細菌学マニュアルにおけるエンテロバクテリア科の家族の説明に対応しており、5つのCSIによって順序の代表者が他のすべての細菌と区別できるという拡大があります。 [初め]
属 Phytobacter チャン et al。 2017 Emend。 ピロンウッド et al。 2018年には家族が割り当てられていません。
以下では、関連する種のほとんどを命名する関連家族のリスト(2019年の時点)。 [6]
- budviciaceae ADORD et al。 2016、fam。 11月。
- ブドヴィシア ブーベット et al。 1985年の修正。 ラング et al。 2013年、Typusgattungそこにある家族
- レミノレラ ヒックマン・ブレナー et al。 1985年
- プラジア ald et al。 1988年

(二次電子顕微鏡記録)
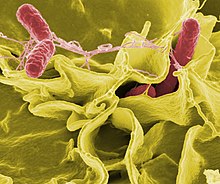


いくつかの同義語と変更 [ 編集 | ソーステキストを編集します ]
- あらゆる種類 レヴィネア 若い et al。 1971年は属になりました Citrobacter ウェルマン&ギレン 1932年に設定
- 違う エルウィニア – タートは属にありました パテアー 、 Enterobacter 、 ペクトバクテリア と ブレンネリア 分割
- LiquidObacterium の同義語です プロテウス
ドイツでは、の直接的な検出です 腸内細菌 特に報告可能 § 7 感染保護法(IFSG)の場合、しかし、カルバペネマーゼ決定剤または自然耐性を除くカルバペネムに対する感度が低下した場合のみです。報告義務は、感染または植民地化の場合にのみ存在します。 (セクション7(1)文1 No. 52 b)IFSG)
さらに、2つ以上の院内感染症の発生を報告する必要はありません。報告する必要はありません。報告は、流行の文脈が可能性があるか疑われる可能性があります。 (() §6 パラグラフ3 IFSG)。
文学 [ 編集 | ソーステキストを編集します ]
- 第6巻:プロテオバクテリア:ガンマサブクラス 。 In:Martin Dworkin、Stanley Falkow、Eugene Rosenberg、Karl-Heinz Schleifer、Erko Stackebrandt(編): 原核生物。細菌の生物学に関するハンドブック 。 3.エディション。 Springer-Verlag、ニューヨーク2006、ISBN 978-0-387-25496-8、doi: 10.1007/0-387-30746-X_9 。
- ドンJ.ブレンナー、J。J。ファーマーIII: 家族I. Enterobacteriaceae 。 In:George M. Garrity、Don J. Brenner、Noel R. Krieg、James T. Staley(ed。): 系統的細菌学のベルゲイマニュアル:第2巻:プロテオバクテリア、パートB:ガンマプロテオバクテリア 。第2版。 Springer-Verlag、ニューヨーク2005、ISBN 978-0-387-95040-2、 S. 587–607 。
- マイケル・T・マディガン、ジョン・M・マーティンコ、ジャック・パーカー: ブロック – ミクロ生物学 、Spectrum Akademischer Verlag、ハイデルベルク、ベルリン2001、ISBN 3-8274-0566-1
個別に [ 編集 | ソーステキストを編集します ]
- ↑ a b c d そうです f g h 私 M. Adeolu、S。Alnajar、S。Naushad、R。S。Gupta: 「Enterobacteriales」のゲノムベースの系統発生と分類:Enterobacterales ordの提案。 11月。 Enterobacteriaceae科、Erwiniaceae Famに分かれた。 11月、Pectobacteriaceae fam。 11月、Yersiniaceae fam。 11月、Hafniaceae Fam。 11月、Morganellaceae Fam。 11月、およびbudviciaceae fam。 11月。 の: 系統的および進化的微生物学の国際ジャーナル。 いいえ。 66 、2016年12月、 S. 5575–5599 、doi: 10.1099/ijsem.0.001485 。
- ↑ Radhey S. Gupta、Emma Griffits: 細菌系統発生における重要な問題。 の: 理論的人口生物学。 いいえ。 六十一 、2002年6月、 S. 423–434 、doi: 10.1006 / tpbi.2002.1589 、 PMID 12167362 。
- ↑ JeanEuzéby、Aidan C. Party: クラスenterobacter。 の: 命名法に立っている原核生物名のリスト、細菌の系統(LPSN)。 2019年12月20日にアクセス 。
- ↑ S. P. Lapage、P。H. A. Sneath、E。F. Lessel、V。D. Skeman、H。P. R. Seeliger、W。A. Clark(Hrsg。)::: バクテリアの国際命名法 – 細菌学コード、1990年の改訂 。 ASM Press、ワシントンDC、米国1992年、ISBN 1-55581-039-X( NCBI本棚 )。
- ↑ P. Fighters、S。P。Glaster、L。K。K. Nilson、T。Hurshansson、L。Guy、S。Guy、S.-J。バス、O。TRENERIUS: Thorsellia Knyensis sp。の提案11月およびThorsellia candunguensis sp。 11月、家族のThorselliae Famのメンバーとして、Arariensisの欠席の最大のものから等しい。 11月 の: 系統的および進化的微生物学の国際ジャーナル。 いいえ。 65 、2015年2月、 S. 444–451 、doi: 10.1099/ijs.0.070292-0 。
- ↑ a b JeanEuzéby、Aidan C. Party: ドメインと門の分類 – 原核生物の階層分類(細菌)。 の: 命名法に立っている原核生物名のリスト、細菌の系統(LPSN)。 2019年12月20日にアクセス 。
Recent Comments