アメリカの先住民の歴史


アメリカの先住民の歴史 これは、染色体の遺伝学やミトコンドリア遺伝学、自己錯性およびタンパク質の遺伝学など、ほぼ同じ物語に収束しているいくつかの分野に基づいています。遺伝的パターンは、アメリカの先住民がいくつかの適切にマークされた遺伝的エピソードを経験したことを示しています。最初で最も重要なのは、シベリアからのアメリカの初期和解であり、古アメリカ人の最初の定住であり、これは現在のアメリンディア人集団に見られる系統と遺伝的マーカーの数の圧倒的要因です。後和解は、北米のNa-denéの人々のそれに対応し、もう1つは北端のスキムローに対応します。それらはすべてシベリアからも。さらに、それは可能です – まだ証明されていませんが、コロンブス以前のアメリカにおけるヨーロッパの遺伝的貢献です。
シベリアの移動の仮説 [ 編集します ]
歴史的背景 [ 編集します ]
人類学的理論は、歴史的にアメリカの先住民をさまざまなグループに置きました。最初の分類(17世紀のフランソワバーニエ)では、彼らはヨーロッパのグループと一緒に分類され、18世紀には独立してグループ化していました。 [ 3 ]
アメリカの先住民がベーリング海峡を通じてアジアから来たことを特に決定したのは、チェコ人類学者のアレシュ・フルドリチカ(1869-1943)でした。 [ 4 ]
3つの移動の仮説 [ 編集します ]
アメリカに人になった3つのシベリア移民理論は、1985年に最初の遺伝的調査で登場しました [ 5 ] そして、1986年以来、言語学者のジョセフ・グリーンバーグ、古政治家クリスティ・ターナー、遺伝学者のスティーブン・ゼグラの作品から、一緒に出版されたことから人気が高まった アメリカの集落:言語、歯科、遺伝的証拠の比較 。 [ 6 ]
言語証拠 [ 編集します ]
ジョセフ・グリーンバーグは、アメリカの言語の3つの主要な家族を提案しました。エスキモ・アルティアナス、Na-dené、Amerindian言語は、アメリカの入植地の3つの異なるプロセスに相当するものです。 [ 7 ] 彼らの方法と結論は、アメリカ人の言語学者の大多数によって受け入れられていませんが(特に、批判は、言語学的観点から系統発生的に有効なグループとしてアメリカのグループを否定することに焦点を当てています)。
最初の2つのグループ(Esquimo-Alutiano、Na-Dené)は普遍的に受け入れられ、最新の2つの波に対応しますが、3番目のグループは、Amerindianが非常に多様であり、最後の2つの移民の場合とは異なり、言語的に多様なグループが参加できたより長い移民プロセスに対応することができます。
これら3つのグループの調査の歴史として、1915年にNa-dené言語がエドワードサピルによって設立されたことが言及できます。 1819年にエスキマレスとaleutasの関係は、19世紀と20世紀の言語学者と人類学者によって受け入れられた1819年にラスマスラスクを決定しました。そして最後に、アメリカ語を明らかに定義している兆候は、1905年にアルフレド・トロンベッティによって述べられ、これは1918年にサピルによって支援されました。
遺伝学と遺伝学は、多種多様な核およびタンパク質鎖を考慮して、数千年および数百万年の分子時計に基づいて結論を得ることができるため、言語的結果と遺伝的結果の比較は相対的です。 [ 8 ] 古石器時代には、現在の証拠に基づいて複数の移動が実施された可能性がありますが、唯一の先祖代々の古アメリカ語のアメリカの仮説を破棄することはありませんが、検証することはできません。 [ 9 ]
物理的人類学的証拠 [ 編集します ]
歯科人類学的証拠は、最初の移民を伴う中国北部の先住民が古アメリカの民族グループの形成を引き起こしたことを示唆した。 Na-denéへの2番目の移行とエスキモエルタの3番目。 [ 十 ]
遺伝分析 [ 編集します ]
血液型 [ 編集します ]
1920年代の研究により、血液型またはコロンブス以前の人口の優位性が決定され、北部でのみ発見されています。ヤコブ・ブロノウスキは、1973年にアメリカの入植地では、少なくとも2つの移動プロセスが与えられると推測しました。最初のグループを運ぶ、または南アメリカの典型であり、北アメリカのようにグループを単独または伴わせる2番目の移動波。 [ 11 ]
タンパク質遺伝学 [ 編集します ]
タンパク質遺伝的研究は、3つの移動の仮説の発展の先駆者です。 1985年の免疫グロブリンGの分析は、ネイティブアメリカンの間で異なる分布を報告しています。これは、シベリア東からのアメリカの入植地に関与する3つのグループに著者に応じて、次のスキームに従って対応しています。 [ 5 ]
| alotipos mg | GM1; 21 | GM1,2; 21 | GM1; 11:13 |
|---|---|---|---|
| プロットマーズ | うん | うん | いいえ |
| Na-den | うん | うん | うん |
| Esquimo-Aleuta | うん | いいえ | うん |
この同じ研究では、GM3ハプロタイプが発見されました。低周波数で5,11,13は、誤認識における白人と可能性のある起源と考えられています。
ミトコンドリア遺伝学 [ 編集します ]
最初に発見された系統はミトコンドリア遺伝学によって与えられ、1990年にアメリカのハプロタイプの4つのグループ(ハプログループ)があります [ 12番目 ] そして、そのうちの1つのバリアントは、NA-DENE村にあります。 [ 13 ] これらの4つのハプログループは、1992年にアルファベットの最初の文字:A、B、C、Dを使用して命名され、アメリカの植民地化のアジア起源をチェックしました。 [ 14 ] ネイティブのNa-DenéDogribDel Canadaでは、グループAによってほぼ排他的に発見されたことを発見しました。これは、AmerindiansがSiberiaからBeringia Bridgeを介して、初期の部族化とともに移住する古い起源を持つため、これがNa-Dené人々の独立した起源を支持するとすぐに推測されました。 [ 15 ]
数年後(1998年)、最初の4つのハプロープA、B、C、Dは東アジアで開発され、西洋のユーロの典型であり、ヨーロッパでヨーロッパにヨーロッパで北アメリカのみの世界で囲まれているため、5番目の系統が発見されました。 [ 16 ]
2014年、メキシコで発見された12 900年のAPで日付が付けられたNAIAのフルスケルトンのミトコンドリアDNA分析は、トゥルムの水中洞窟のシステムであり、古アメリカ人と現代アメリカ人の間の遺伝的リンクをテストしました。 [ 17 ]
これらの結果は、シベリアからのアメリカの入植地の理論を支持しています。 [ 18 ] ルートとそのおおよその古代は、次の地図に示されています。

染色体と [ 編集します ]
アメリカの先住民には、1995年に決定された明らかに過半数の父系系統が1つしかありません。それはDYS199(現在はQ-M3またはQ1A2A1A1)と呼ばれ、エスキモスを含むすべてのアメリカの先住民族で提示されていますが、特に中央アメリカと南アメリカでは90%以上の頻度があります。 [ 19 ]
その後、他の系統が決定され、特に北米ではホパログループCとRが決定されたため、シベリアからの2つの主要な移動がある可能性があると推測されました。 [ 20 ] 次の分布を生み出します。
Q-M3(Q1A2A1A1、Q3以前)はアメリカ全土で広まっていますが、その存在は特に最初の移動、古アメリカ人の移動に関連しています。その後の移動は、他の系統をもたらし、したがって、ハプログループC-P39(C2B1A1A、以前のC3B1)とEskimos Q-NWT01(Q1A1A)が優勢であるNa-dené村では、優先されました。 [ 22 ]
常染色体遺伝学 [ 編集します ]
ヌクレオチド多型で実施された最新の遺伝学的研究は、アメリカの沈降は約15、000年前に最初のアメリカ人が到着し、その後Na-denéとEsquimoaleutianの人々が到着したアジアからの3つの移動波で行われたと結論付けています。 [ 23 ]
メイングループ [ 編集します ]
その後、アメリカのネイティブ集団で行われた地域の線形遺伝子分析(ハプログループ)は、特定の母性元の遺伝的コレクション(ミトコンドリア)と父方(染色体および)がそれらを定義できるグループとサブグループを特定できるようにします。
次の表では、他の人々との誤認識に起因する可能性のある可能性のある括弧内(北から南への秩序)が大胆にあります。
| グループ | Subrupo | 母体の系統 | 父方の系統 |
|---|---|---|---|
| Esquimo-Aleuta | エスキモー | A2a、 A2B 、D2、D3 | Q-NWT01 、Q-M3(R1)。 |
| アリュータ | A2a、 D2 | Q-M3( R1 )。 | |
| Na-den | Na – デネデルノート | A2A (D2)。 | Q-M3、C-P49(R1)。 |
| Na-DenéDelSur | A2A (B2、C1)。 | Q-M3 、C-P49 | |
| プロットマーズ | 北米 | A2-64 、B2、C1、D1、X2 | Q-M3(C-P49、R1)。 |
| メソアメリカと南アメリカ | A2-64、B2、C1、D1、D4H | Q-M3 |
類似点と相違点 [ 編集します ]
アメリカの先住民の間では、違いよりも類似点があります。常染色体の遺伝学によると、アメリカ大陸では遺伝的多様性が低く、代わりに世界の天然集団と大きな差別化があります。 [ 24 ] この単位と遺伝的均一性の証拠は、アメリカの和解A2(ADNMT)とQ-M3(DNA-Y)の設立系統が大陸の典型であり、しばしば支配的であることです。血液型または+のように。
最も顕著な違いは、連続した移民波と新しい系統の導入との関係の結果です。したがって、旧石器時代の間、最初の古系アメリカ人の系統は、a、b、c、d1(adnmt)、q-m3(dna-y)、そしておそらくx(dnamt)の後に到着していたでしょう。一方、新石器時代のC(DNA-Y)は、おそらくNa-denéの人々と、d2、d3(adnmt)およびq-nwt01(dna-y)がエスキモスと一緒に来ました。
アメリカの集落に関する最後のより詳細な研究、 [ 25 ] 52のアメリカの村、17のシベリア人、300,000のDNAシーケンスを分析します。 [ 26 ]
- アメリカのネイティブ人口の大半は、ベリンギアが渡った最初の移民の単一グループ、氷河期の間にアジアとアメリカを結びつける橋から完全に降りています。そこから大陸全体に拡大があり、その後複数の発散が続き、最終的にはアメリカのネイティブグループ、特に南アメリカではほとんど遺伝的流れがありませんでした。
- Na-denéとEsquimouleutasの言語のスピーカーの間に遺伝的フットプリントを残した2つの後続の移行がありました。これらの移動は、すでに確立された最初の先住民と混ざり合っており、Na-denéの人々は最初の移動のゲノムの大部分である90%を呈し、その50%が保存されていました。
- エスキモ・シベリアの人々の遺伝的分析とチュクチスで見られるように、アメリカからシベリアへの移行もありました。
Paleoamericanos [ 編集します ]

古石器時代(10、000年以上前)にアメリカに到着した最も古い集団と呼ばれ、多くの遺伝学者によってアメリカインド人とも呼ばれています。
染色体分析では、メソアメリカと南アメリカで100%の周波数でハプログループQを見つけることが一般的であるため、古アメリカの遺伝的遺産の一部であったに違いありません。同じ現象は、ラテンアメリカの多くの民族グループでグループまたは100%の頻度である血液型分析で発生します。 [ 27 ]
古序的遺伝学 [ 編集します ]
現在の先住民集団のDNA分析と人間の化石の分析は、アメリカの入植地の研究に新しい光を提供します。 7000年前(世界で最も古い)までのチンチャロミイラの42のサンプルの分析は、31.2 b:21.9 c:31.2およびd:3.1%ミトコンドリアプロファイルをもたらします。 [ 28 ]
アラスカでは10 300年の人間の遺体が発見され、ミトコンドリアハプロープD4H3および染色体Q-M3に属していることが報告されました。 [ 29 ] しかし、最も古い遺伝的証拠は、オレゴン州(米国)のキューバス・デ・ペイズリーの12 300年のヒトコプロライトの分析であり、アメリカの古 – アメリカの起源のプロビスをサポートし、A2とB2(ADNMT)をもたらしました。 [ 30 ]
古代 [ 編集します ]
アメリカ大陸への最初の入植者の到着は、彼らの出身国と年齢の両方で広く議論されており、12、000年または最大60、000年とは異なる日付を提案しています。
最近の研究は、この植民地化がSo -Called Called Ultimate Glacial Maximumの後に発生したことに同意しています。この研究では、主要なパンアメリカンハプロープA2、B2、C1、およびD1に基づいて19,000歳の年齢を示しています。 [ 最初に30 ] そして、アラスカのA2(ADNMT)の起源の可能性に基づくもう1つは、シベリアからのアメリカの植民地化に17、000年を与えます。 [ 32 ] 属プロジェクトは、15、000年から20、000年前に発生したと述べています。 [ 33 ]
染色体の遺伝学とアメリカの植民地化によると、それは約14、000年になるでしょう、 [ 21 ] 自営業の遺伝学によると、彼は約15,000歳です [ 25 ] そして、すべてのアメリカのミトコンドリアグループの分子時計の分析によれば、それは約15,000歳の年齢であり、これは考古学的登録の最も信頼できるソースと一致します。 [ 34 ]
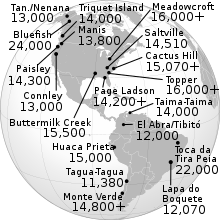
最も信頼できる考古学的証拠は、最も古いものなどの認識された遺物は、約14、700年のモンテヴェルデのものであり、1300年のクロービス100よりも古いことを確認しています。 [ 35 ] 特に南アメリカでのはるかに古いデートに関する報告は、出会い系の方法や手順の批判のために科学界の多くに認識されておらず、論争が維持されています。 [ 36 ]
このようにして、近年発表されたすべての遺伝的証拠は、20、000年以前の可能性の低い初期の和解に対する後期人口理論を支持しています。大陸全体の植民地化は速く、北米と南アメリカの極端な間の約2000年の程度の差がありました。 [ 37 ]
シベリアのつながり [ 編集します ]
シベリアからのアメリカの集落は広く認識されています。なぜなら、アメリカとアジアの氷河期の間、彼らはベリンギアと呼ばれる大きな橋を通して結びついていたからです。 [ 38 ]
遺伝的証拠は、ネイティブアメリカンをシベリアの先住民に関連付けるため、この論文を強化します。 4つのパン – アメリカのハプロープA、B、C、およびD(ADNMT)は、シベリア南部全体で十分に拡張されており、モキエーターと持っています。たとえば、Tuvanosでは、4つのグループが72%を追加し、最初のアメリカ人とシベリア南部の間に可能性のあるつながりを確立することにより提示されます。 [ 39 ] しかし、AとBが希少なシベリア北部では、ハプログループCとDがYakutosとEnvenkis Orientalで86%を追加するようになります。 [ 40 ]
一方、すべてのアメリカの先住民族の間で最も一般的な父系系統Q(DNA-Y)は、ケットやセルクップなどの10年前の村の西シベリアで最大の頻度を持っています。 [ 41 ]
結論として、シベリアとアメリカの間にはつながりがあるが、現在、ユーラシアにはアメリカの入植地と絶対に関係する特定の地域はないと言えます。 [ 39 ]
シベリアの先祖の起源 [ 編集します ]
アメリカに順番に住んでいた最初のシベリア移民は二重起源を持っていた、つまり、彼らは白人男性とモンゴルの女性の子孫であったと考えられています。これらの特徴を持つ人々は、今日シベリア中央に生息しています。 [ 42 ]
24、000年の南シベリアの子供のゲノム分析は、この二重性を裏付けています。東アジアと西ユーラシアの人口の誤解は、アメリカ先住民の先祖のコレクションの一部でした。 [ 43 ]
古アメリカの植民地化ルートの仮説 [ 編集します ]

最初の入植者の渡りルートに関するいくつかの仮説があります。地質学的証拠に従って最も受け入れられているのは、の仮説クローブです 氷のない廊下 最後の氷河作用の終わりに、ベリンギアと北米の大平原との間に形成されていたでしょう。太平洋の沿岸沿岸のルートも可能です。
最後に、ヨーロッパからの移住の可能性が維持されています、 ソルトレンス仮説、 アメリカ人の祖先の相続の一部として。ただし、遺伝的証拠に基づいて、これらの3つのルートのいずれも完全に実証されていませんが、破棄されていません。
単一の移行仮説 [ 編集します ]
ミトコンドリアの証拠によると、古アメリカの植民地化は、沿岸ルート上のシベリアからの単一の移動の結果である可能性があります。 [ 44 ]
仮説2シベリアからのルート [ 編集します ]
シベリアからのアメリカの2つの古石器時代のコロニー形成ルートのアイデアは、古系アメリカ人が染色体YまたはミトコンドリアDNAまたは両方によって、2つのルート、太平洋岸に沿った非常に古いルート、および2番目の沿岸に沿って2つのルートを生成することによって、2つの遺伝的に分化したサブグループに合理的に分割できるという事実に由来しています。 [ 45 ] [ 46 ] さまざまな著者に従って次のように要約できる2つの集団を特徴付ける氷のない廊下によって:
| グループ | 可能性のあるルート [ 9 ] | 古代 [ 35 ] | 母体の系統 [ 9 ] | 父方の系統 [ 21 ] |
|---|---|---|---|---|
| パン – アメリカのアメリカ人 | 太平洋沿岸の移動 | + 17 500 | A2、B2、C1、C4C、D1、D4H3A [ 9 ] | Q |
| 北米東部のアメリカインディアン | 氷のない廊下 | -12 600 [ 46 ] | A2、B2、C1、D1、X2A、X2G [ 9 ] | Q(C3B、R1B)。 |
これらの理論は、10 300年前からプリンスオブウェールズ島(アラスカパシフィックコスタ)でハプログループD4H(D10とも呼ばれる)の発見によって強化されています。 [ 47 ] 一方、ケネウィックの男はQ-M3(DNA-Y)およびX2A(ADNMT)ハプログループを紹介しました。 [ 48 ]
北米東:西系統 [ 編集します ]
北米東部のハプログループX(DNAMT)の存在 (マップを参照) 、西ユーラシアの典型的な系統は、いくつかのDNAの存在と一致しており、R1Bや他の未成年者などの西部ユーラシアの典型的なもの(G、J、E)も一致しています。 [ 49 ] これらの西部系統のアメリカでの存在は、次の3つの仮説で解釈されています。
シベリア起源 [ 編集します ]
アメリカにおけるx(adnmt)、r1a、r1b、c(dna-y)の存在は、シベリアから氷のない廊下を介した移動の結果である可能性があります。 [ 35 ] これらのすべての系統が見つかる可能性があるため、シベリアの南部と中心部の現在の集団ではほとんどほとんどありません。 [ 50 ] [ 51 ] [ 52 ]
この仮説には、先住民族の民族的および文化的な一致があります。 [ 20 ]
最近のヨーロッパの誤解 [ 編集します ]
アメリカのR1Bと他のDNAと未成年者の存在は、アメリカ人とヨーロッパ人の間の現代の誤認識の結果であると考えられています。 [ 49 ] この仮説は、オジブワの人々とヨーロッパの商人との間の接触の間の民族誌的証拠と一致するでしょう、 [ 53 ] 母体の系統は、主にカナダの先住民とアングロフランスの植民地化の父方の植民地化から来るメチスと同様の高い誤解を生み出します。 [ 54 ] [ 55 ] 他の著者はこの仮説を支持していますが、 [ 21 ] [ 56 ] 他の仮説を排除できることを可能にする詳細なサブクラディスティック分析は、実行または公開されていません。
ソルトレンス仮説 [ 編集します ]
他の著者は、特にイベリアからのヨーロッパからの移住のためのアメリカの植民地化の可能性を維持しており、15、000年前にはハプログループXにもたらされ、ソリューションとクロービス文化の間のつながりを構成します。 [ 57 ] Xはヨーロッパ全体、特にアイルランドで一般的であり、X2J系統が発見されました。これはアメリカの系統X2Aの布兄弟です。 [ 58 ] さらに、R1bは西ヨーロッパで支配的です。 [ 59 ] 一部の著者は、岩の産業クロービスを、ツールの製造のスタイルに基づいて、ソルトレンス文化のクロビと同一視しています。 [ 60 ] しかし、決定的な証拠があるとは考えられていません。 [ 六十一 ] また、約7500歳でヨーロッパ人に似たケネウィックの男性の遺体は、アメリカの入植地の初期段階でのヨーロッパの移民に関連している可能性があることも示唆されています。 [ 62 ]
また、10世紀以来のアメリカのバイキング植民地化に関連している可能性もあり、ヴィンランドのネイティブスクレリングはカナダのアルゴンキンの人々への言及である可能性があります。 [ 63 ] [ 六十四 ] または、北欧やケルトの村では、これらのハプログループ(x2-adnmtおよびr1b-adn-y)が存在するため、アルゴンキーノとセルタの関係がある可能性があります。
Amerindianサブグループ [ 編集します ]
現在、アメリカの人口がサブグループに分割できるようにする遺伝的分類は認識されていません。ただし、常染色体およびミトコンドリアの遺伝学により、言語的側面や地理的側面と一致するいくつかの関係を確立することができますが、これらのグループの明確な数を定義したり、それらの間の明確な境界を確立することはありません。
|
常染色体遺伝学 [ 編集します ]1988年のCavalli-Spforと彼のチームによる研究、 [ 65 ] 遺伝的多型について世界中の42の集団を調べて、いくつかのマクロパミリアの遺伝学と言語学の間に相関関係があると結論付け、アメリカ原住民の3つの大規模なグループと北アジア(シベリア)の集団(左のクラドを参照)も関連しています。 |

分子遺伝学研究所、イギリス、スイス、コロンビア、ペルー、メキシコ、カナダ、ブラジル、コスタリカ、チリの分子遺伝学研究所の間の常染色体および調整遺伝学の最近の研究(2007年)は、世界の残りの24人のアメリカ人の個体群を54人の米国先住民族と関連付けています。 [ 24 ] アメリカ大陸内では、遺伝的多様性が低く、代わりに世界のネイティブ集団と大きな差別化があると結論付けています。結果の中で (正しい画像を参照) あなたは、アメリカの人々(チペワン)とアメリカ人との間の最大の相違を見ることができます。そして、アメリカの中で、北米東部の人々は、一方では、他方(メキシコから南アメリカの極端なものまで)によって、他方によってアメリカの人々とアメリカ人の人々と区別することができます。ケチュアとアイマラの間の密接な遺伝的関係も観察できます。
ミトコンドリアのサブグループと言語ファミリー [ 編集します ]

さまざまな地理的領域は、大型ハプロープA、B、C、D、Xの分布の形式と相関しており、これは最も認識されている言語ファミリーに対応しています。次の地域は(南から北へ)最も代表的なものです。
- アラウカニアとパタゴニア: センターとチリ南部とアルゼンチンの人々は、ハプログループの優位性によって特徴付けられています d 、続いてCが続いて、Mapuches、Yaganes、AlacalufesなどのSo -Calcled South Andean言語のスピーカーがそれに調整されています [ 六十七 ] Y選択。 [ 68 ]
- 南アメリカからの東: アマゾン地域の人々によって形成された不均一なグループ、グランチャコ、カリブ海、アンティル諸島を含む。 4つのパン – アメリカのハプログループは、系統が少し際立っていますが、それらのどれも明らかに優勢ではありません。 c 、たとえば、アララウアの家族、カリブ海、トゥカナ、ヤノマミ、パノタカナ、 [ 72 ] ティクナ、コファン、モビマ。 Mataco-GuaicurúグループのDへの部分的な傾向がある、 [ 71 ] あなたがたのBに向かって、GuaraníesのAに向かって。 [ 七十三 ]
- ノースアメリンディオ: ハプログラポ a 北米、米国北部、カナダの東部で過半数です。これは、アルギック、イロコイ、シー、ムスコグイの特徴です [ 76 ] サリシャノスとワカッシュ。ただし、この地域の最も特定はハプログループの存在です バツ 、それは均一な分布を持っていません。なぜなら、遠くから最大の頻度と多様性があるため、分布は均一ではありません。 [ 44 ] Xの分布とA-Dセットの分布の違いは、これらの人々の二重起源の可能な証拠として解釈できます。Xは後に初期集団に吸収されたでしょう。 [ 9 ]
プエブロとデネ [ 編集します ]

さまざまな遺伝学的研究では、スピーカーがエスキモエレタとアメリカの人々のNA-デネ語と区別しています。しかし、Na-denéの人々を最も特徴づけるのは、Hoplogroup C3(DNA-Y)の存在です。 [ 77 ] (マップを参照)
Na-denédelsur y del Northe [ 編集します ]
米国南西部のタンパク質遺伝的研究(Alotipo Mg)は、この地域で最も重要な遺伝的違いは、南Na-Dené(Apache、Navajo)と他の人々(Pima、Pápago、Hopi、Hualapai)の中にあることを発見しました。 [ 5 ]
14の遺伝子遺伝子座からの研究では、アタバスカナのポップリケーション(Na-dené)は、アルゴンキンの人々よりもエスキマレとチュクチスに近いことがわかりました。 [ 78 ] [ 79 ] 後部cavalli-sforza(1994) [ 80 ] 彼は、23人のアメリカの民族グループを分析するより徹底的な研究を行いました。その結果、彼は北の北の人々から南の人々から距離があることを確認し、エスキモスに最も近い北の人々のように見え、代わりに南ナ・デネがアミャノス・アモサノスの人々(Ietic-Wakash)に近づくだろう。
ミトコンドリアDNAを考慮して、HASAを含む北Na-denéはHapogroup A(Adnmt)の絶対的な優位性を示していますが、南Na-Denéには、北米のアメリンディアンの人々との誤認識が発生したと判断できる他のマイナーなグループがあります。 [ 81 ]
Yenise Connection [ 編集します ]

言語の肯定は、言語と言語の言語における言語学を紹介し、デネ・エンセアの家族に適合します。 [ 82 ]
染色体の遺伝的証拠と、これらの人々の間のつながりも与えてくれます。ケットシベリアの人々は、実際には元の言語を保持している唯一のイェニウスの町であり、2つのハプログループのみによって形成されたADNとプロファイルを持つ唯一のものです。過半数QとC3マイノリティ、 [ 41 ] Apacheや他のNa-denéの人々のように。
ミトコンドリア遺伝学に関しては、A2A1母体系統は、アパッチ(Na-dené)とセルクピス(イェニセオス)の両方で見つかりました。 [ 29 ]
Esquimo-Aleutas [ 編集します ]
彼らはエスキモ・アルティア語の話者であり、 スケール 。 [ 83 ]
ミトコンドリアの遺伝学は、特徴的なハプログループは2500年から5000年前の比較的最近のものであるため、他のアメリカのネイティブグループに関して最新の起源であると判断しました。最も古い遺跡はグリーンランドから来ており、約4000年前であり、遺伝的に分析されており、約5500年前に最初のSki-On-Landがシベリアから到着したと信じることができます。 [ 84 ]
Origen dual [ 編集します ]
特徴的な系統スケールは、D2、D3、A2A、A2B(ADNMT)およびQ(DNA-Y)です。シベリアから来た移動は彼にハプログループD2とD3をもたらしましたが、A2とそのA2AおよびA2Bのサブシラーはアメリカ起源であると考えられています。 [ 29 ] したがって、スケールタスの人々は二重起源を持っているでしょう。一方では、シベリアから、もう一方は、最後の氷河作用の終わりに氷に戻った後に北極圏に植民地化したネイティブアメリカンのもう一方では。これが、エスキマレにQ-M3(DNA-Y)が存在する理由である可能性があります。
チュクチ接続 [ 編集します ]
同様に、チュクチの人々は、これらの同じハプログループD2、D3、A2A、およびA2Bを紹介します。 [ 32 ] これらの人々の間の密接な遺伝的関係のシグナル。これはまた、アメリカに住んでいるシベリアの移住だけでなく、シベリアに戻ったアメリカの移住が彼らの系統のいくつかを運んでいることを意味します(A2A、A2B、およびおそらくC1A)。 [ 29 ] 次の表を使用すると、この関係を観察できます。
| グループ | 母体の系統 [ 32 ] | 父方の系統 |
|---|---|---|
| イヌイット | A2a、 A2B 。D3 | Q-NWT01 、Q-M3、R1 [ 22 ] |
| aleutas | A2a、 D2 | Q-M3( R1 )) [ 85 ] |
| シベリア語 | A2A 、A2B、D2、D3 | N1C 、Q-M3、R1 [ 20 ] |
| チュクチス | A2A 、A2B、C2、C3、D2、D3、G | C3C、 N1C 、Q-M3、R1 |
Esquimo-EleutasとChukchisの間のこの遺伝的なつながりは、Esquimo-Alutian言語とChucoto-Kamchaktas言語(Swadesh 1962)との関係において言語的同等物を持つ可能性があります。 [ 八十六 ] または、ウラロシベリア語の仮説にさえ関係しています。
系統の古代 [ 編集します ]
ミトコンドリアDNAによると [ 編集します ]
DNAの変異の数が多いほど各ハプログループの年齢が大きいという仮説に基づいて、各系統の古さを計算しようとしました。ミトコンドリア分子時計に基づく2つの最近の研究 [ 34 ] [ 八十七 ] 次のおおよその結果を示します。

- a :約30、000年前にオリエンタルアジアで生まれました。
- A2以前(またはA4):25、000年および中国起源の可能性があります。
- A2:13 000 [ 34 ] または、アラスカで可能性が高い16、000年 [ 32 ]
- A2A:4700、北米と東シベリア。 [ 32 ]
- A2B:2600、Esquimals and Chukchis。
- A2(64):A2D、A2G、A2H(コロンビア)、A2I(USA)、A2J、A2K、A2N(カナダ)、A2P(エクアドル)、A2R(メキシコとグアテマラ)など、アメリカでサブクロールされたいくつかのサブクロールがあります。
- A2:13 000 [ 34 ] または、アラスカで可能性が高い16、000年 [ 32 ]
- A2以前(またはA4):25、000年および中国起源の可能性があります。
- b (16189、8281)東洋アジアでは50、000年。
- B4:44,000
- B4B:28 000
- B2 :14 600では、アメリカ原産です。
- B4B:28 000
- B4:44,000
- M8 :43、000年、東端の典型。
- c :28 000、典型的なシベリア。
- C1:17 000
- C1A:13 900、アジア
- C1B:14 500、アメリカ
- C1C:9000、アメリカ
- C1D:19 000で15,000、アメリカで [ 88 ]
- C4:18,000
- C1:17 000
- c :28 000、典型的なシベリア。
- d (16189、8281)48、000年、東極端の典型。
- D4:27,000
- D1:13 500、アメリカから排他的。
- D4E:23 000
- D4E1:19 000
- D2:13、000年、シベリアの南で発生しました。 [87]
- D2a: 5000 años y con la frecuencia más alta en aleutas. Propio de esquimales y chukchis.[32] Encontrado en Saqqaq (Groenlandia), en restos de hace 4000 años.[89]
- D2:13、000年、シベリアの南で発生しました。 [87]
- D4E1:19 000
- D3(O D4B):26 000
- D3A(O D4B1):20 000。
- D3A2A(またはD4B1A2A):シベリアとエスキマレス。
- D3A(O D4B1):20 000。
- D4H:18 000、東アジアおよびアメリカで。 [ 9 ]
- D4H3B:アメリカの典型。彼は12、600年前にクロービス文化に身を包んだ。 [ 90 ]
- D4:27,000
- バツ (16189、8281)32、000年で、近東で始まりました。
- X2:21 000
- x2a’j:北米に加えて、イラン、シベリア(ポドカメンナヤ・トゥングスカ)でも発見されました。 [ 58 ] アイルランドとエジプト。
- X2A:12 800年、北米からの排他的。
- x2a’j:北米に加えて、イラン、シベリア(ポドカメンナヤ・トゥングスカ)でも発見されました。 [ 58 ] アイルランドとエジプト。
- X2:21 000
染色体によると古代 [ 編集します ]
ネイティブアメリカンの最も一般的な系統であるQ、C、およびRでは、これらのおおよその古代は以下で計算されます。
- c :約50、000年前にアジア南部に生まれました。 [ 77 ]
- C2:イーストエンドから所有しています。
- C-P39:14、000年、 [ 49 ] プエブロのデネ(アメリカ人のアメリカ)のはしかを綴ります。
- C2:イーストエンドから所有しています。
- p :南アジアで生まれ、約34、000年前のものです。 [ 91 ]
- Q :南アジアで最大の多様性。
- Q1A1(F1096):近東および極東で拡張。
- Q1A1A*(NWT01):イヌイットの村で、4000〜7000年の支配者。 [ 22 ]
- Q1A2(M346):ユーラシア、特に中央アジアとシベリアで拡張。
- Q1A2A1(L54)アメリカおよびヨーロッパ人。
- Q-L54*(XM3)は、Q-M3後にアメリカインディアンの中で最も頻繁なグループまたは段落です。彼は12、600年前にクロービス文化に身を包んだ。 [90]
- Q1A2A1A1(M3):典型的なAmerindio、13、000年 [49]または多分22,000。 [92]アラスカで300年前の10人の人間の遺体で発見されました。 [47]
- Q1A2A1(L54)アメリカおよびヨーロッパ人。
- Q1A1(F1096):近東および極東で拡張。
- r :27 000 [ 91 ]
- R1B:西の典型である18、500年前に近東で生まれました。
- Q :南アジアで最大の多様性。
参照してください [ 編集します ]
参照 [ 編集します ]
- ↑ モンゴロイドの形成 Mapa de las migraciones humanas 、九州博物館のウェブサイトで。
- ↑ Burenhult、Göran(2000)。 最初の人々 。 weltbild verlag。
- ↑ レッスン、レーネプライムバレ(1827)。 哺乳類のマニュアル、またはマミファーの自然史 。
- ↑ Hrdlicka、A。(1942)。 アメリカインディアンの起源と古代 。ワシントン。
- ↑ a b c ウィリアムズ、R。C。、および。アル。 (1985)。 «ネイティブアメリカンのGMアロタイプ:ベーリングランドブリッジを横切る3つの明確な移動の証拠»。
- ↑ グリーンバーグ;ターナー; Y Zegura(1986)。 «アメリカ大陸の和解:言語、歯科、および遺伝的証拠の比較»。
- ↑ グリーンバーグ、ジョセフ(1987)。 «アメリカの言語»。
- ↑ Cavalli-Sforza、Luigi Luca(1992)。 «遺伝子、人、言語»。
- ↑ a b c d そうです f g 前に、ugo、そして。アル。 (2009)。 «2つの珍しいmtDNAハプログループでマークされたベリンギアからの独特の古インドの移動ルート»。 の: 現在の生物学 19、2009年1月1日、p。 1-8。
- ↑ ターナー、C。G。(1986)。 «太平洋のリム集団の髄relolocalsology分離推定»。
- ↑ ブロノウスキー、ジェイコブ(1975)。 男の上昇 。 ロンドン:BBC(Broadcasting Corporation)。ページ。 92-9 ISBN 0 563 10498
- ↑ Schurr、T。G。、et。で。 (1990)。 «アメリンインディアンミトコンドリアDNAは、高周波数でまれなアジアの突然変異を持ち、4つの主要な母体系統に由来することを示唆しています»。
- ↑ トロニ、アントニオ、他に。 (1991)。 «ネイティブアメリカンのミトコンドリアDNA分析は、AmerindとNadeneの個体群が2つの独立した移動によって設立されたことを示しています»。
- ↑ ウォレス、ダグラスC。 Y Torroni、Antonio(1992)。 «ミトコンドリアDNAに書かれているアメリカインディアンの先史時代:レビュー»。
- ↑ トロニ、アントニオ、他に。 (1993a)。 «Aderindiosの4つの設立グループのアジアの親和性と大陸放射»。 アーカイブ 2006年9月24日、ウェイバックマシンで。
- ↑ a b ブラウン、マイケル、他アル。 (1998)。 «mtdna haplogroup X:ヨーロッパ/西アジアと北米の間の古代のつながり?»。
- ↑
- ↑ 「北アジア、アメリカ人のゆりかご」 アーカイブ 2014年5月17日Wayback Machineで。 ユカタンの北 (2014年5月16日)。
- ↑ アンダーヒル、ピーター、他アル。 (1995)。 «コロンブス以前のY染色体特異的移行と人間の進化史への影響»。
- ↑ a b c レル、ジェフリー、他うなぎ。 (2001-2002)。 «ネイティブアメリカンの二重起源とシベリアの親和性»。
- ↑ a b c d ボルトロニ、メアリー・カンティラ、他午前。 (2003)。 «y-染色体は、アメリカ大陸における古代の人口統計学的歴史が異なることの証拠»。 アーカイブ 2011年4月28日、Wayback Machineで。
- ↑ a b c Dulik、Matthew C。; Owings、A。C。; Gaieski、J。B。; Vilar、M。G。;アンドレ、A。;レニー、C。;マッケンジー、M。A。; Kritsch、I。; Snowsho、S。;ライト、R。;マーティン、J。;ギブソン、N。;アンドリュース、S。D。; Schur、T。G.(2012)。 «Y染色体分析は、アタパスカンおよびエスキモン語を話す集団における遺伝的発散と新しい設立ネイティブ系統を明らかにします»。 の pnas 、109(22):p。 8471-8476。
- ↑ «アメリカの和解に関する最も詳細な遺伝的研究»。 アーカイブ 2012年7月17日、Wayback Machineで。毎日 トレード (ペルー)、2012年7月。
- ↑ a b Wang、Sijia、et。al。2007、 ネイティブアメリカンの遺伝的変異と人口構造
- ↑ a b ライヒ、デビッド、他に。 2012年 ネイティブアメリカンの人口歴史の再構築 Nature 2012 doi:doi:10.1038/nature11258
- ↑ «ネイティブアメリカンの人口は、3つの重要な移動から降りてくると科学者は言います»。 雑誌 科学毎日 、2012年7月。
- ↑ 啓発された教訓的な百科事典 、ボリュームIX。スペイン:サルバト、1987年。
- ↑ a b モラガ、マウリシオ、他で。 (2001)。 «チリ北部のミイラにおけるミトコンドリアDNA分析は、アンデスの人口のアマゾン仮説をサポートしています»。
- ↑ a b c d そうです タム、エリカ、他アル。 (2007)。 «ベーリングの停止とネイティブアメリカンの創設者の拡散»。
- ↑ ギルバート、トーマス、他に。 (2008)。 «北米オレゴン州のクロビス以前のヒトコプロライトからのDNA»。 雑誌 化学 、9、vol。 320、n.º5877、págs。 786-789、2008年5月。doi:10.1126/science.1154116
- ↑ アキレス、アレッサンドロ、および。アル。 (2008)。 «4つの汎アメリカのmtDNAハプログループの系統発生:進化および疾患研究への影響»。
- ↑ a b c d そうです f Volodko、Natalia、et。アル。 2008、 北極シベリアのミトコンドリアゲノムの多様性。特に、アメリカ大陸のベリンギアの進化的歴史とプレイストセニックの人々が言及しています
- ↑ 人間の旅/遺伝マーカーのアトラス。 の: ナショナル・ジオグラフィック 。
- ↑ a b c ソアレス、ペドロ、他で。 (2009)。 «選択の補正の修正:改善されたヒトミトコンドリア分子時計»。 そして彼のページ 補足。 の: American Journal of Human Genetics 、第84巻、N.O 6、ページ。 740-759、6月4日、
- ↑ a b c シュール、セオドア; Yシェリー、スティーブン(2004)。 «ミトコンドリアDNAおよびY染色体の多様性とアメリカの人々の人々:進化的および人口統計的証拠»。
- ↑ ディレヘイ、トーマス(2000)。 «最初のアメリカ人の考古学を議論する»。 アーカイブ 2014年12月14日、Wayback Machineで。の第2章 アメリカ大陸の定住 。
- ↑ Kumar、Satish、et。アル。 (2011)。 «メキシコ系アメリカ人における大規模なミトコンドリアシーケンスは、ネイティブアメリカンの起源の再評価を示唆しています」。 の: BMC進化生物学 、11:págo。 293、doi:1186/1471-2148-11-2
- ↑ Alllcious John f。;エリアス、スコットA.(2007年6月15日)。 ベリンギアの人間生態学 。コロンビア大学出版局。 p。 3. ISBN 978-0-231-13060-8。
- ↑ a b Derenko y Malyarchuk 2001、 ネイティブアメリカンの祖先を探して (アメリカのアボリジニの先祖の家を探して)
- ↑ デレンコ、ミロスラバ、他アル。 (2007)。 «北アジアの個体群におけるミトコンドリアDNAの系統地理学的分析»。
- ↑ a b タンベット、クリスティナ、それ。アル。 (2004) «サアミの西部と東部の根 – ミトコンドリアDNAとY染色体によって語られた遺伝的な「外れ値」の物語» ; American Journal of Human Genetics 74 :661–682。
- ↑ »先住民はシベリアからアメリカに到着しました»。 の: コスタリカのロシアの官報 、2010年。
- ↑ ウェイド、ニコラス2013。 24、000年前の体は、ヨーロッパ人とアメリカのインディアンに親族関係を示しています ニューヨークタイムズ
- ↑ a b Bagundes、N。、および。アル。 (2008)。 «ミトコンドリアの集団ゲノミクスは、アメリカの人々のための海岸沿いのルートを持つ単一のクロビスの起源をサポートしています» (PDF) 。 American Journal of Human Genetics 82 (3):583-592。から提出 オリジナル 2009年3月25日 。 2009年11月19日に取得 。
- ↑ Callaway、Ewen(2016年8月10日)。 «植物と動物のDNAは、最初のアメリカ人が沿岸ルートをとったことを示唆しています» 。 自然 536 (7615) 。 2016年8月23日に取得 。
- ↑ a b ペダーセン、ミケル。ウィラースレブ、エスケ、et.al。 (10 de Agosto de 2016)。 «北米の氷のない廊下における氷河後の生存率とコロニー形成» 。 自然 。 2016年8月23日に取得 。
- ↑ a b ケンプ、ブライアン、他に。 (2007)。 «アラスカからの初期の完新世の骨格残遺物の遺伝的分析とアメリカ大陸の和解への影響»。
- ↑ Morten Rasmussen et al。 2015、 ケネウィックマンの祖先と所属 Nature(2015)doi:10.1038/nature14625
- ↑ a b c d Zegura、Stephen、他で。 (2004)。 «高解像度のSNPとマイクロサテライトハプロタイプは、アメリカのネイティブアメリカンY染色体の単一の最近のエントリをアメリカに指し示しています»。
- ↑ Khar’kov、V。N.、et。アル。 (2009)、 «Y染色体マーカーデータから推測されたテレウトの遺伝子プールの比較特性»。 アーカイブ 2011年8月5日、Wayback Machineで。
- ↑ Derenko、M.、et。に。 (2001)、 «南シベリアのアルタイ人におけるミトコンドリアハプロープxの存在»。 の American Journal of Human Genetics 、69(1):p。 237-241。
- ↑ Reidla、Maere; Kivisild、T。; Metspalu、E。アル。 (2003)。 「MtDNA Haplofroup Xの起源と拡散」。 en: American Journal of Human Genetics 、73(5):p。 1178-1190。
- ↑ ロードス、R。(1982)。 «ガルゴンキン貿易言語»。 en:Cowan、W。(編集者): 第13回アルゴンキア会議の議事録 。オタワ:カールトン大学。 PGS。 1-10。
- ↑ ««北西部のメティスネーション»。» 。から提出 オリジナル 2012年2月26日 。 2011年7月31日に取得 。
- ↑ ««ojibwe History»。» 。から提出 オリジナル 2013年1月24日 。 2012年10月29日に取得 。
- ↑ Mhalhi、Ripan Singh、et。アル。 (2008)。 «ネイティブ北米人の間のY染色体の分布:アタパスカの人口歴史の研究»。
- ↑ ウォレス、ダグラス。 «ネイティブアメリカンハプログループ:ヨーロッパの系統»。 ドーランDNAラーニングセンター。
- ↑ a b Reidla、Maere; Kivisild、T。; Metspalu、E.、et al。 (2003)。 «mtDNAハプログループxの起源と拡散»。 の: American Journal of Human Genetics 73(5):p。 1178-1190。
- ↑ Mc Donald、J。D。(2005)。 ハプログループマップ。 アーカイブ 2013年12月11日、Wayback Machineで。
- ↑ ブラッドリー、B。; Yスタンフォード、D。(2006)。 «ソルトリアクロビス接続»。
- ↑ Carbonell、Eduard(2006)。 人類:大陸の最初の職業 (P. 627)。マドリード:アリエル。
- ↑ ustrate、glynn。 «ケネウィックの男の禁じられた発見»
- ↑ «L’Anse Aux Meadowsでバイキングを発見します»。 ニューファンドランドとラブラドールの遠隔学習とイノベーションセンター、カナダ。
- ↑ Wahlgren、Erik(1990)。 バイキングとアメリカ 。バルセロナ:目的地。 84-233-1915-6 。
- ↑ Cavalli-Sforza、L。L.、et。に。 (1988)。 «人間の進化の再構築:遺伝的、考古学的、言語データをまとめる»。
- ↑ «相互接続された世界地域における生きている人間の新しい遺伝的地図»。 アーカイブ 2011年9月29日、Wayback Machineで。 DNA部族。
- ↑ ガルシア、フェデリコ、他で。 (2004)。 «チロ諸島の集団の起源と微小拡散化»。 (インターネットアーカイブで利用可能な壊れたリンク。 記録 、 最初のバージョン そしてその 最後 )。
- ↑ ソテー、セルジオ、他で。 (2010)。 «エスクエルの遺伝子混合物と片側系統(チューバット州)。他の人口サンプルとの比較»。 (インターネットアーカイブで利用可能な壊れたリンク。 記録 、 最初のバージョン そしてその 最後 )。
- ↑ Sandoval、ホセ、他で。 (2004)。 «湖の島のチチカカの島民のADNMTバリアント:B1ハプロタイプの最大周波数と設立効果の証拠»。 (インターネットアーカイブで利用可能な壊れたリンク。 記録 、 最初のバージョン そしてその 最後 )。
- ↑ a b c メルトン、フィリップ、他アル。 (2007)。 «中央および南アメリカのチブチャンの話す人口の生物学的関係:mtDNAからの証拠»。
- ↑ a b Citlalin、Xochime(2002)。 «赤茶色の地球の人々»。
- ↑ イーストン、ルース、他アル。 (1996)。 «ヤノマミのmtDNAバリエーション:追加の新しい世界設立系統の証拠»。
- ↑ サラ、アンドレア、他に。 (2010)。 «核およびミトコンドリアの多型マーカーによる北東アルゼンチンに住むムビア・グアラニの6つのコミュニティの遺伝的分析»。
- ↑ すぐに、eduvigis(2006)。 «ヒスパニック前から植民地の中mesoまで:古いDNAのフットプリント»。
- ↑ マリ、リパン、他アル。 (2003)。 «アメリカ南西部のネイティブアメリカンmtdna先史時代»。
- ↑ 患者、デボラ、他アル。 (2003)。 «米国南東部のネイティブアメリカンの間のミトコンドリアDNA変動の予期しないパターン»。
- ↑ a b 「Hoplogroup C」分散。 の: ナショナル・ジオグラフィック 。
- ↑ Szathmary、E。J。E.(1981)。 «シベリアおよび北米北部の人口の遺伝的マーカー»。 en 年鑑物理人類 、24:p。 37-73。
- ↑ Szathmary、E。(1985)。 «北米の人々:遺伝的研究からの手がかり»。 en:Kirk、R。; Szathmary E.(編)。 «アジアから:アメリカと太平洋の人々の人々。 en Pacific Historyのジャーナル 。キャンベラ、p。 79-104。
- ↑ Cavalli-Sforza、Luigi Luca(1994)。 «人間の遺伝子の歴史と地理»。
- ↑ mtdna«Amerind Founder Haplogroup Project -Goal»。 In:ftdnaプロジェクト。
- ↑ Vajda、Edward(2008)。 «na-dene言語とのシベリアのリンク»。
- ↑ ダモンド、ドンE.(1965)。 «Eskaleutian言語学、考古学、および先史時代»。
- ↑ ラスムッセン、モーテン、他うなぎ。 (2009)。 «絶滅したPalaeo-eskimoの古代のヒトゲノム配列»。
- ↑ Rubicz、Celeste(2007)。 «司令官とプリビロフ島で最近設立されたアレウツコミュニティの進化的結果»。
- ↑ スワデシュ、モリス(1962)。 «Behring Strait»を介した言語関係»。 en aa 、64:p。 1262-1291。
- ↑ a b Derenko、M.、et。に。 (2010)。 北アジアのミトコンドリアDNAハプロープCおよびDの起源と後氷河分散
- ↑ 前に、ugo、そして。アル。 (2010)。 «アメリカ大陸の最初の人々:ベリンギアからの創立ミトコンドリアゲノムの増加»。
- ↑ ギルバート、トーマス、他に。 (2008)。 «Paleo-eskimo mtDNAゲノムは、グリーンランドのマトリニールの不連続性を明らかにします»。
- ↑ a b Rasmussen M、Anzick SL et al。 2014、 モンタナ西部のクロービス埋葬地からの更新世後期の人間のゲノム。 自然。 2014年2月13日; 506(7487):225-9。 doi:10.1038/nature13025。
- ↑ a b カラフェット、タチアナ、他アル。 (2008)。 «新しいバイナリの多型は、人間のY染色体ハプロープツリーの解像度を再構築および増加させます»。
- ↑ ビアンキ、ネスター、他で。 (1997)。 «DNA分析によるアメリカの識別»。 (インターネットアーカイブで利用可能な壊れたリンク。 記録 、 最初のバージョン そしてその 最後 )。
外部リンク [ 編集します ]
Recent Comments