ミトコンドリアエヴァ – ウィキペディア

ミトコンドリアEVA は考古学からの用語であり、ミトコンドリアDNA(mtDNA)からの女性を示しています。あなたの男性のカウンターパートは、Y染色体のアダムです。
異なる個人の異なる遺伝子(またはゲノムの他の分化したセクション)を見ると、通常、異なる個人にはわずかに異なるバリアントがあることがわかります(ゲノムの多型)。個人が対応している、短い相同遺伝子またはDNAセクションである場合、これらのバリアントは、元の配列が突然変異によって徐々に変化したという事実からのみ生じる可能性があります。これはすでに相同遺伝子の定義に対応しています。したがって、それぞれの(ホモロジスト)DNAセクションは、今日の多様性が徐々に発達した元のシーケンスに精神的にまでさかのぼることができます。時間プロセスでこのプロセスに従うと、それぞれが突然変異している分割プロセスのパターンが生じます。代わりに、今日のシーケンスを撮影して起源を再構築しようとすると、振り返ってみると、これらのそれぞれが「フロー」に対応しています(英語」 合体) それぞれのシーケンス。したがって、対応する分析は、合体分析と呼ばれます。
人間では、すべての染色体、したがって各遺伝子は、原則として2つのコピーで、1つは母親から、もう1つは父親(ディプロイド)から来ています。性染色体に加えて、例外はミトコンドリアの独立した遺伝子です。これらは、主に細胞のエネルギーサプライヤー(「発電所」)として機能するオルガネラです。精子の特別な建物のため、すべてのミトコンドリアは、男性と女性の両方で、卵細胞から来て、母体のゲノムを運びます。各個人のミトコンドリア遺伝子は、すべて同じハプロタイプを持っています。ミトコンドリアDNAの合体分析により、今日の変動幅を起因するDNA配列を計算できます。この特定のシーケンスの着用者は、やや印象的で、「ミトコンドリアEVA」と呼ばれます。
この祖先自体の存在は些細なことであり、特別な知識とは関係がない – それは、すべての人々、実際にはすべての生物が最終的に関連しているため、ある時点で共通の祖先を持っていたに違いないという事実から自動的に生じる。論理に関しては、種の形成はすでにかなりの多型を持つ可能性のある集団に衰退しているため、「ミトコンドリアEVA」は私たちの種類に属してすらないはずです。そのため、ミトコンドリアのシーケンスが生じる共通の祖先はすでに前身から来ることです。 [初め] すでに多型と変異速度は異なる遺伝子で異なりますが、単に他の遺伝子を見るとき、単に偶然からも、「ミトコンドリアのエヴァ」よりもはるかに若いまたは古い異なる共通の祖先もあります。雇用は、この些細な基本的な声明を、たとえば分割の場所や時間にさらなる知識に追加できる場合にのみ科学的に興味深いものになります。
理想的な条件(無限の人口サイズ、無制限の混合、適応等しい対立遺伝子、変異なし)では、集団に存在するすべての対立遺伝子は、完全に変化しない周波数(Hardy-Weinberg平衡)で長期的に保存されます。これは、実際の集団では決してそうではありません。人口規模が限られているため、対立遺伝子は、個人の異なる子孫によって、ランダムによってより頻繁にかかるか、それほど一般的になります。この現象はジャンルと呼ばれます。その結果、ジャンルによるアレレンの寿命は、たとえそれらが完全に同等(中立的)であっても、実際の集団では限られています。人口サイズが小さくなるほど、より強くなければならないことがわかります。ランダムパス(数学的にはMarkowチェーン)は、開始サイズに関係なく、1つの左(アレルの「固定」と呼ばれる)のみが他の変異の存在下で、より遅いか遅れて集団内のすべてのアレルを導きます。半数体遺伝の場合(ミトコンドリウムと同様)、前世代1の合体の確率は、人口サイズn(実際には異なる交配確率によって個人の数に加えて影響を受ける「有効な」人口サイズ)で分割されます。これにより、人口規模の2倍の合体時間の期待値が得られます。 [2] 人口規模は100人で、共同祖先が約200世代になると予想されます。ただし、実際の集団では、新しい対立遺伝子を生成する突然変異の影響を考慮する必要があります。
ミトコンドリアEVA 過去の特定の時期に最初の女性でも唯一の女性でもありませんでした。 エヴァ 多くの同時代人がいました。しかし、他の女性のミトコンドリアの相続は死にましたが、 エヴァ 生き残った(多かれ少なかれ多くの突然変異を持つ子孫の大部分で)。この祖先の生涯と場所は、今日住んでいる代表的な数の個人のmtDNAの助けを借りて、正確に絞り込むことができます。
多くの特性により、mtDNAは人間の降下を研究するための貴重なツールになります。
- 細胞核のDNAと比較して、mtDNAはより高い一定の突然変異率を示します。
- mtDNAは母親によってのみ渡されるため、有効な人口サイズは常染色体DNAのサイズのみであり、両方の親がそれぞれ2つのコピーを持っています。したがって、対立遺伝子の固定は約4倍速いです。したがって、ヒトmtDNAの対立遺伝子は、常染色体DNAよりもはるかに若く、ユーラシアの定住などの若い人類の歴史を研究するのに非常に適しています。
- 変異速度とmtDNAは、常染色体DNAと比較して高く、対立遺伝子の頻度が部分集団から他の集団に変動することを意味します。これらの違いから、集団の降下、移動、変位、または混合に関する声明は、地理的に均質な常染色体DNAよりもはるかに簡単に導き出すことができます。
- 細胞には多くのミトコンドリアが含まれており、それぞれにmtDNAのいくつかのコピーがあるため、化石(例:ネアンデルタル骨組織)は分析のために抽出されることがよくありますが、細胞コアDNAははるかに頻繁に引き渡されます。
- mtDNAの継承における組換えの欠如は、女性の相続線の特定の特性に関する声明を可能にします。

ヒトミトコンドリアDNAの変動の初期研究は、1983年にすでに実施されました。 [3] [4] この理論は、レベッカ・L・カン、アラン・ウィルソン、マーク・ストーンキング(1987)による出版で知られるようになりました。 [5]
can et al。 (1987)世界のさまざまな地域の女性の胎盤からmtDNAを抽出しました。それらはmtDNAを配列しませんでしたが、制限フラグメント長多型(RFLP)を使用して調査を実施しました。彼らは類似性に応じて家系図にmtdnasを注文し、最終的に家系図の根を決定しました。 2つの主要な枝は、決定された根から分岐しました。アフリカ人だけが、世界のすべての地域の他の人々に自分自身を見つけました。このことから、著者はと結論付けました ミトコンドリアEVA アフリカに住んでいたに違いありません。
彼らはまた、分子時計を使用して、いつ ミトコンドリアEVA 住んでいます。 1987年には、魚、鳥、およびいくつかの哺乳類種のミトコンドリアDNA(mtDNA)に関するデータがすでにありました。このデータは、mtDNAが100万年あたり約2〜4%変化したことを示しました。このデータは、分子時計の校正に使用されました。研究のヒトミトコンドリアDNAは平均で0.57%のみ異なっていたため、それは結果をもたらしました。 ミトコンドリアEVA ちょうど約200、000年前に住んでいたに違いありません。
著者によると、拡大する人口における相続の固定はありそうもないので、議論はしなければなりません ミトコンドリアEVA 今日住んでいるすべての人々の祖先がアフリカを去る前に住んでいました。著者のアフリカ外理論の明確な兆候。 「現代人の多地域起源」の仮説は、著者によって拒否されました。 ミトコンドリアEVA これのためにはるかに古いはずです( 立っている男の人 ほぼ200万年前にアフリカを去っていた)。

この出版物は最初からひどく批判されました。多くの批判は正当化されたように見えた:
- この研究の147人のうち、20人の「アフリカ人」のうち2人だけがサハラの南アフリカから来たのは2人だけでした。他の18人はアフリカ系アメリカ人でした。
- 家系図を生成する方法は、必ずしも統計的に最も安いツリーを提供するわけではありませんでした。
- ツリーの根を見つけるために、根は最も長い枝の中央に配置されました( ミッドポイントルート化 )。これは、ルートの間違った位置につながる可能性があります。 B.アフリカの進化速度が高い場合。
- RFLPは、分子時計にとって重要な突然変異率を決定するのにあまり適していません。
- 悪い統計分析。
したがって、(深刻な)批判は一般的にの概念に反するものではありませんでした ミトコンドリエンエヴァ 指示。この問題に精通している人は皆、この女性がどこかにどこかに住んでいたに違いないことは明らかでした。著者の科学的アプローチのみが批判されました。
しかし、後で、改善された研究が確認され、Cannからの最も重要な声明を支えました et al。 (1987)。たとえば、IngmanがLED et al。 (2000)新しい改善された研究を通じて:
- 彼らは53人のリハーサルを受け、そのうち32人はサハラの南アフリカのさまざまな地域からリハーサルをしました。
- 彼らはMTDNA全体を押収しましたが、分析のために彼らはすぐにそれらを閉じました 進化 Dループ領域オフ。
- 家系図の根は、チンパンジーのmtDNAを使用して決定されました( アウトグループのルート化 )。
この改善された研究の結果は、1987年の研究よりもさらに明確でした。
- アフリカ人と非アフリカ人の完全な分離。
- 家系図の最初の3つの枝はアフリカ人にのみつながり、4番目はアフリカ人と非アフリカ人につながりました。
- アフリカの長い枝ですが、外側の星の形の構造(最近の拡張の特徴)。
- 共通の祖先まで175,000±50、000年 ミトコンドリアEVA 研究のすべての人々)。
- 最後のアフリカと非アフリカの支部の間で分岐するために52,000±28、000年 ミトコンドリアEVA 研究のすべての非アフリカ人)。
- 約1925年のアフリカの支部での拡張のシグナル、つまり20年の世代を想定している場合、約38、500年前。
2013年がついに登場しました 化学 99,000〜148、000年前の「ミトコンドリアEVA」と、120,000〜156、000年前のいわゆるY染色体のアダムによると、別の研究が発表されました。 [6]

数字は、変異の位置を示しています。
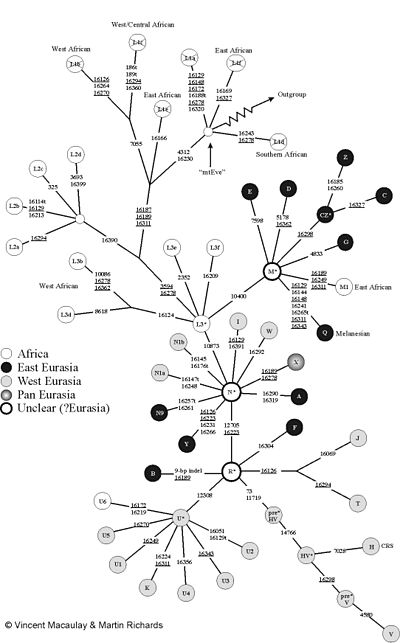
数字は、変異の位置を示しています。
「mtev」はそれです ミトコンドリアEVA 。 「アウトグループ」は、他の霊長類(チンパンジーなど)のmtDNAにつながります。図は、「L1ハプロープ」:L1を使用して通常の(偽の)命名法を使用しますが、ルートを形成します(L1AはV!よりもL1Fとは関係ありません)。したがって、L1フィールドは消去されました。 [7]
人間のミトコンドリアDNAは、SO -Called Haplropsに分けることができます。次に、ハプリンググループには他のサブハプロップを含めることができ、それをさらに細分化できます。このツリー構造をハプロップの命名法にマッピングしようとし、文字と数字を交互に使用します。ハプログループの2つのMTDNAは常に単系統です。割り当てでは、Dループの外側のmtDNAの遺伝子配列で特徴的な変異が使用されます。
一人がzをできます。 B.ハプログループC1A3B2を持っています。彼女のmtDNAは、zの他の人と密接に関連しています。 B. C1A3B4。もちろん、彼女のmtDNAはC1A3C5を持っている第三者と共通の祖先を共有していますが、この共通の祖先はC1A3ラインが分裂する前に住んでいました。これは、C1A3B4およびC1A3B2がC1A3C5と比較して単系統であることを意味します。 C1A3B2およびC1A3C5は、すべてのH-Haplotypesなどと比較して単系統です。
残念ながら、命名法は比較的矛盾しています。多くの文字が、最も重要な非アフリカのハプロップに名前を付けるために使用されました。ただし、多くの古いHAPLグループがアフリカで発生しています。これは一緒に「L」と呼ばれ、メイングループの分割のためにすでに数字に転送されています。現在までに、いくつかのアフリカのハプロタイプの割り当てについて科学的なコンセンサスはまだありません(L1およびL3)。
ルートから始めると、男性のミトコンドリアの家系図は、最初は多くの深い枝で構成されています。これらの遺伝線は今日L1と呼ばれています。かつてのこととは反対に、L1は単系統ハプログループではなく、ルートを形成します。したがって、L1は実際にはそれと同じくらい古いアフリカのハプロップの全体のパッケージです ミトコンドリアEVA そして、お互いの正確な関係はまだ明らかにされていません。
これらの古いL1枝のうち、枝は10810の位置の突然変異によって分岐します。次に、ハプログループL2は、16390の位置の突然変異によってこれから分割されます。 L2は、サハラの南のアフリカ人のみでのみ実質的に発生します。
位置3594の突然変異が枝を形成し、その上で大きなハプルッペMとN、および他の多くのアフリカのハプロップがあり、現在もL3の下で要約されています。 L3は、L1と同様に、実際の(単系統)ハプログループではありません。 haplruppe mとnは、非アフリカの大多数で発生します。サハラの南アフリカでは非常にまれで、L1、L2、L3が支配しています。
ハプログループMは、大きなハプロップM1、Z、C、D、E、G、Qに分割されます。 N1A、N1B、N9、A、I、W、X、Y、およびサブハプログループB、F、H、P、T、J、U、およびKを形成するハプログループRのハプログループN。
ミトコンドリアDNAの現在最も広範な検査はそうでした ジェノグラフィックコンソーシアム 実行された(a。theGenographic Projectを参照)。この比較では、78,590個の遺伝子型サンプルが含まれ、ミトコンドリアのハプロップ(およびそのサブグループ)が系統樹に示されました。 [8]
地理的分布 [ 編集 | ソーステキストを編集します ]
L枝の「古い」ハプロタイプは、サハラの南アフリカで支配的です。あなたがそこにあなたの起源を持っていることは間違いありません。これらのハプロタイプは、北アフリカ(約50%の頻度)およびヨーロッパおよび西アジアでも、低頻度でも見られます。
Haplruppe MとNは、世界の他の地域で支配的であり、サハラの南アフリカではまれです。ハプログループM(M1)の特別なバリアントは、エチオピアで約20%の頻度で発生します。どちらかがすでにそこに作成されているか、それはセムの南翼のハイキングです。
Haplruppe A、B、C、D、およびXは、アメリカの先住民の間で発生します。一方、Haploruppe N、CおよびDの東部枝からのA、B、およびXは、Haplogroup Mから作成されました。
ヨーロッパと西アジアでは、ハプログループは非常にまれです。最も一般的なサブグループは、サブグループR:H、V、T、J、U、Kに属します。さらに、ハプログループI、W、Xも有意な周波数で発生します。ヨーロッパ、コーカサス、および中東では、個々のハプロップの周波数のみを変動させるのとほぼ同じハプロップがあります。特に、ハプログループHは、中東およびコーカサスではヨーロッパよりもはるかに一般的ではありません(約25%のVers約45%)が、ハプログループKは大幅に一般的です。地域に応じて、HAPLグループの周波数はヨーロッパ内で変動します。
南アジアと東アジアは、ハプロルチェンの西アジアとは大きく異なります。ここでは、ハプログループMから、ハプログループC、D、E、G、Z、Q。 Haploruppe Nもここで発生しますが、主にHaplogroup A、B、F、Y、Xによって表されます。
Haplistruppe Xは、比較的低い頻度でのみユーラシアと北米全体で発生するため、注目に値します。過去には、Haplistruppe Xがヨーロッパで作成され、ヨーロッパでのみ発生すると想定されていました。アメリカ先住民の間でハプリンググループが発見されたとき、ヨーロッパ間の海でヨーロッパからアメリカに来たという仮説が生じました。それまでの間、Haplistruppe Xもアジアで発見されました(Derneko et al。 、2001)。
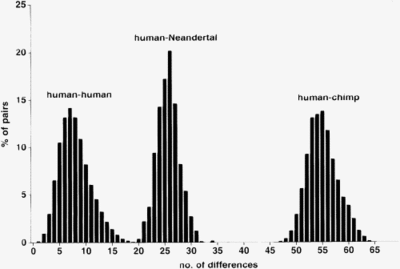
各細胞には多くのミトコンドリアが含まれており、ミトコンドリアDNAのいくつかのコピーが含まれています。これにより、多くの場合、化石のmtDNAのDNA配列決定が可能になります。 2006年まで、MtDNAの一部は、ドイツ、クロアチア、ロシア、フランス、ベルギー、イタリア、スペインの12人のネアンデルタール人によってシーケンスされていました。 [9] 結果:
- ネアンデルタール人のmtDNAは、2人の現代人のmtDNAの約3倍異なります。
- ネアンデルタール人のmtdnaはほとんど異なります。現代の人々のmtdnaはもっと異なります。
- ネアンデルタール人のmtDNAは、今日のアフリカ人やアジア人のヨーロッパ人よりもムトナとの類似性を示していません。
ネアンデルタール人のmtdnaは、現代の人間につながった線から比較的早い段階で分裂しました。 1つは、共通点を推定しています ミトコンドリアEVA 現代人とネアンデルタール人から550,000〜690、000年前、つまりそれよりもはるかに早い ミトコンドリアEVA 現代人の(クリングス et al。 、1997)。
Nordborg(1998)などによれば、このデータにもかかわらず、ネアンデルタール人と現代人の間に混合があることを除外することはできません。一方、Currat&Excoffier(2004)は、このmtDNAデータは実際には除外できると述べました。しかし、2012年から2014年までの研究結果は、ルーマニアのペシュテラ・キュー・オアシスの化石と、ヴァスト・イジムがネアンデルタール人とのハイブリダイゼーションを実証しました。 [十] [11]
すべての化石からのmtDNAは解剖学的に現代の人々のままです( ホモサピエンス )、これまでに検討されてきたが、今日の生活者と密接に関連している(Serre et al。 2004)。たとえば、「Ötzi」のmtDNA et al。 2006)、より正確にはサブタイプK1F。 [12番目]
ミトコンドリアDNA配列の多様性は、アフリカで最大です。全体として、人間の遺伝的多様性は、類人猿と比較して低いです。 MtDNAの特定のセグメントでは、チンパンジーは人と比較して3〜4倍の遺伝的多様性を示しています。 19のMTDNAは、タイ国立公園(コートジボワール)からチンパンジーを調べましたが、これらのサルは小さな遺伝的混合グループに属しますが、すべての人々の多様性よりも高い多様性を示しています(Gagneux et al。、 1999)。
ボノボス、ゴリラ、オランウータンも、人間よりもMtDNAの遺伝的多様性が高いことを示しています(ジョブリング et al。、 2004)。
| 軌跡 | チンパンジー vs.メンシュ |
ボノボ vs.メンシュ |
ゴリラ vs.メンシュ |
オランウータン vs.メンシュ |
参照 | 注釈 |
|---|---|---|---|---|---|---|
| mtdna | 3〜4倍高い | より高い | より高い | より高い | 勝者 et al。 (1999) | – |
| Y染色体 | より高い | より高い | 以下 | より高い | 結石 et al。 (2002) | アダムを参照してください |
| X-染色体 | 3倍高い | データなし | 2倍高い | 3.5倍の高さ | Kaessmann et al。 (2001) | – |
| 軌跡 | アフリカ | アジア | ヨーロッパ | 参照 | 注釈 |
|---|---|---|---|---|---|
| mtdna(ペアの違い) | 2.08 | 1.75 | 1.08 | 警戒 et al。 (1991) | – |
| Y染色体(43マーカー) | 0.841 | 0.904 | 0.852 | ハンマー et al。 (2001) | アダムを参照してください |
| X-染色体(ヌクレオチドの多様性) | 0.035 | 0.025 | 0.034 | Kaessmann et al。 (1999) | – |
| 常染色体(ヌクレオチドの多様性) | 0.115 | 0.061 | 0.064 | yu et al。 (2002) | – |

の理論 ミトコンドリエンエヴァ 現在、認識され確立されていますが、アフリカ外の理論の問題と「現代人の多領域起源」の問題に関しては、科学的なコンセンサスはまだありません。の理論が明らかであることは明らかです ミトコンドリエンエヴァ アフリカ外の理論と矛盾していません。しかし、それは必ずしも他のモデルに反論するわけではありません。結局のところ、彼女は単一の遺伝子座についてのみ声明を出します。細胞核内の遺伝子座には、組換えのおかげで他の物語があります。
たとえば、Nordborg(1998)は、Neanderthal-Mtdnaの完全な不在が、今日のヨーロッパ人の間のGendriftの結果である可能性があると主張しました。
Nordborg(1998)は、単純なモデルで、現代人が最初はネアンデルタール人と混合せずにヨーロッパに広がったと仮定しました。拡張が完了して初めて、現代人はネアンデルタール人と混ざり合った。このモデルによると、ネアンデルタール人は、それを作った遺伝子プールに最大25%の寄与を持っていたかもしれません。それまでの間、ネアンデルタール人のミトコンドリアの継承はその間に死亡しました(その間に 参照してください 仮説)。今日のヨーロッパ人は、ネアンデルタールのurprungを含む細胞核にまだDNAを持っている可能性があります。
Currat&Excoffier(2004)は、現代の人間とネアンデルタール人を混合するためのはるかに複雑なモデルを開発しました。ヨーロッパの和解と現代の人々によるネアンデルタール人の移動は、約12、000年(約500世代)に続きました。拡張とミキシングが同時に期限切れになったという計算を想定した場合、ネアンデルタール人は事前にヨーロッパに住んでいたため、大きな利点をもたらします。
zの場合。 B.アフリカから来た現代人、アナトリア大佐、時にはネアンデルタール人と混ざり合って、MtDNAは現代の人間の遺伝子プールへのネアンデルタール人の入り口を見つけます。現代人の人口は現在拡大しており、輸入されたネアンデルタール語のDNAも拡大しています。今zの場合。 B.バルカン人には「混合子孫」が住んでおり、そこに住んでいるネアンデルタール人との混合が再びあり、遺伝子プールのネアンデルタール語のDNAの割合は増加し続けています。現代人の拡大の波が西と北の拡大の波がさらに浸透し、常にそこに住んでいるネアンデルタール人と混ざり合うほど、より多くのネアンデルタール語のDNAは遺伝子プールで豊かになります。
このモデルによれば、小さな混合クォータでさえ十分であるため、ネアンデルタール人のミトコンドリアDNAは現代の人間のミトコンドリアDNAを完全に置き換えます(そうではありませんでした)。 Currat and Excoffier(2004)は、現代人とネアンデルタール人がヨーロッパに住んでいた12、000年で、最大120人の「混合子供」しか存在できないと判断しました。この非常に少数のため、著者は、現代人とネアンデルタール人が一緒に再現できない異なるタイプであると疑っていました。
アジアの大部分はすでに属の代表者から来ています ホモ ( 立っている男の人 )彼らがアフリカから現代の人々が住んでいたときに住んでいた。ここでも、混合を示すミトコンドリアの継承はありません。
要約すると、それはと言えます ミトコンドリアEVA アフリカ外の理論は、次の事実をサポートしています。
- ヒトのミトコンドリアDNAは、遺伝的多様性が低い(Gagnenuxなど、1999)。
- ミトコンドリアEVA わずか175,000歳で比較的若い年齢(Ingman and Others、2000)があります。
- ミトコンドリアの家系図は、アフリカの深い枝を示していますが、外の星の形の構造を示しています(Ingman and others、2000)。
- ネアンデルタール人のミトコンドリアDNAは、今日の人々とは明らかに異なります(Serre and others、2004)。
- ネアンデルタール人との現代人の混合は、2004年までありそうもないと考えられていました(Currat&Excoffier、2004)。
X染色体、Y染色体および常染色体に関する他の遺伝子座に関する研究では、若いアフリカ人の人間の起源も示しています(Takahata and others、2001)。
2013年から2015年まで、スウェーデンの科学者SvantePääbo周辺の研究チーム 混合 公開:
- 改善された分析方法は、今日のヨーロッパ人およびアジア人の遺伝子プールに最大4%のネアンデルタール人遺伝子の寄与により、遺伝子の流れが起こったことを示しました。 [13]
- の研究データ マンサピエンス – ルーマニアのペシュテラ・キュ・オアシスとシベリアのヴァスト・イディシムによる化石は、これらの声明を支えました。 [十] [11]
- ただし、これまでのところ、このフローは一方向でのみ証明されています。 マンサピエンス – ネアンデルタール人女性と男性。 [14]
- ブライアン・サイクスには呼ばれる本があります 7人の娘エヴァス 書かれています。
- エデンの川で、リチャード・ドーキンスは遺伝子から川の文脈で人間の祖先を説明し、 ミトコンドリアEVA 多くの一般的な祖先の1つは、さまざまな遺伝的経路に基づいて追跡できることです。
- ディスカバリーチャンネルには、ドキュメンタリーがあります 本当のエヴァ ブロードキャスト。
- 日本の小説、ホラー映画、ビデオゲームシリーズのパラサイトイブは、致命的な結果をもたらす再生細胞の助けを借りて妻を復活させる科学者に関するファンタジーの物語の基礎として、ミトコンドリアのエヴァ理論を使用しています。
- グレッグ・イーガンはタイトルで短編小説を書きました ミトコンドリアEVA 。
- ロナルドD.ムーアズサイエンスフィクション – スリーズバトルスターギャラクティカIST ヘラ 人の娘とカイロニンとして ミトコンドリアEVA 。
- リン・オカモトは、彼の漫画のためにミトコンドリアのエヴァの理論を使用しました 11曲 。これで、ルーシーはディクロニウスと呼ばれる新しい品種のミトコンドリアのエヴァです。
- 専門文学
- マーカスラン、L。エクセッフィア: 現代の人間は、ヨーロッパへの範囲の拡大中にネアンデルタール人と混ざりませんでした。 の: PLOS生物学。 ローレンス2.2004,12、E421。 doi: 10.1371/journal.pbio.0020421 ISSN 1544-9173 。
- M.V. Derenko、T。Grzybowski、B.A。 Malyarchuk、J。Czarny、D.M。 Sliwka、I.A。ザハロフ: 南シベリアのアルタイア人におけるミトコンドリアハプロープXの存在。 の: American Journal of Human Genetics (Am J Hum Genet)。ニューヨーク69.2001、1、237-241。 PMID 11410843 ISSN 0002-9297 。
- P.ガグヌー、C。ウィルズ、U。ゲルロフ、D。タウツ、P.A。モリン、C。ボーシュ、B。フルース、G。ホーマン、O.A。ライダー、D.S。ウッドラフ: ミトコンドリアシーケンスは、アフリカのヒノイドの多様な進化の歴史を示しています。 の: アメリカ合衆国科学アカデミーの議事録 (PNAS U S A)。ワシントン96.1999,9、5077-5082。 doi: 10.1073/pnas.96.9.5077 ISSN 0027-8424 。
- M.F.ハンマー、T.M。 Karafet、A.J。 Redd、H。Jarjanazi、S。Santachiara-Benerecetti、H。Soodyall、S.L。 ZEGURA: グローバルなヒトY染色体多様性の階層パターン。 の: 分子生物学と進化 (Mol Biol Evol)。オックスフォード18.2001、7、1189–1203。 PMID 11420360 ISSN 0737-4038 。
- M. Ingman、H。Kaessmann、S。Pääbo、U。Gyllennen: ミトコンドリアのゲノム変異と現代人の起源。 の: 自然。 ロンドン408.2000,6813、708-713。二: 10.1038/35047064 ISSN 0028-0836 。
- マーク・A・ジョブズ、クリス・タイラー・スミス、マシュー・ハルズ: 人間の進化遺伝学。起源、人々、病気。 ISBN 0-8153-4185-7。
- H. Kaessmann、F。Heissig、A。VonHaeseler、S。Pääbo: ヒトX染色体上の低組換えの非コーディング領域におけるDNA配列変動。 の: 自然遺伝学 (Nat Genet)。ニューヨーク22.1999,1、78–81。 doi: 10.1038/8785 ISSN 1061-4036 。
- H. Kaessmann、V。Wiebe、G。Weiss、S。Pääbo: 類人猿DNAシーケンスは、多様性の低下と人間の拡大を明らかにしています。 の: 自然遺伝学 (Nat Genet)。ニューヨーク27.2001、2、155–156。 doi: 10.1038/84773 ISSN 1061-4036 。
- M.キングス、A。ストーン、R.W。 Schmitz、H。Krainitzki、M。Stoneking、S。Pääbo: ネアンデルタルDNA配列と現代人の起源。 の: 細胞。 ケンブリッジ90.1997、1、19-30。 doi: 10.1016/s0092-8674(00)80310-4 ISSN 0092-8674 。
- M.ノードボルグ: ネアンデルタール人の祖先の確率について。 の: American Journal of Human Genetics (Am J Hum Genet)。ニューヨーク63.1998、4、1237-1240。 PMID 9758610 ISSN 0002-9297 。
- F.ロロ、L。ヘルミニ、S。ルシアニ-IIIオリビエリ、D。ルイゼリ: アイスマンのmtDNAハプログループの細かい特性。 の: American Journal of Physical Anthropology (Am J Phys Anthropol)。ニューヨーク130.2006、4、557-564。 doi: 10,1002/aver。 ISSN 0002-9483 。
- Serre、D。; Langunyy、A。;頬、M。;ニコラ、M.T。; Paunovic、M。; Menencier、P。; Hefreiser、m。;ファンには、G。&Pubeboes、S。(2004): 初期の現代人へのneandertal mtdnaの貢献の証拠はありません。 PLOS Biol 2(3)、E57。 doi: 10.1371/journal.pbio.0020057 。
- ストーン、A。C。;グリフィス、R。C。; Zegura、S。L。&Hammer、M。F。(2002): PAN属の高レベルのY染色体ヌクレオチド多様性。 Proc Natl Acad Sci U S A 99(1)、43–48。 doi: 10.1073/pnas.012364999 。
- 徹夜、L。; Stoneking、M。; Harpending、H。;ホークス、K。&ウィルソン、A。C。(1991): アフリカの個体群と人間のミトコンドリアDNAの進化。 Science 253(5027)、1503–1507。 doi: 10.1126/science.1840702 。
- タカハタ、N。;リー、S.H。 &Satta、Y。(2001): 現代の人間の起源の多面性のテスト。 Mol Biol Evol 18(2)、172-183。 PMID 11158376 。
- あなた、いいえ。チェーン、フィン。 .tter、SM、SM。羊、u。パムロロ、P。 thethy、y。ブラット、m。 Turs、tes。あなたが見る、S。&2002):(202): アフリカ人とユーラシア人の間では、アフリカ人の中でより大きな遺伝的違い。 Genetics 161(1)、269–274。 PMID 12019240 。
- 受信
- ブライアンサイクス: 7人の娘エヴァス。 Lübbe-verlag、Bergisch Gladbach 2001、ISBN 3-7857-2060-2。
- ↑ J.ハイン、M.H。 Schierup、C。Wiuf: 遺伝子系図、変動、進化:合体理論の入門書。 オックスフォード大学出版局、オックスフォード2005。
- ↑ アラン・R・テンプルトン: ハプロタイプの木と現代の人間の起源。 物理人類学の年鑑48、2005、S。33–59。
- ↑ C.F.アクアドロ、B.D。グリーンバーグ: ヒトミトコンドリアのDNA変異と進化、7人の個人からのヌクレオチド配列の分析。 (PDF; 1.7 MB) の: 遺伝学。 jg。 103、Bethesda 1983、S。287–312。 PMID 6299878 ISSN 0016-6731
- ↑ M.J.ジョンソンu。 a。: 制限エンドヌクレアーゼ切断パターンによって分析されたヒトミトコンドリアDNAタイプの放射。 の: Journal of Molecular Evolution。 jg。 19、ニューヨーク1983、S。255–271。 PMID 6310133 doi: 10.1007/BF02099973 ISSN 0022-2844
- ↑ R.L. Cann U.A: ミトコンドリアDNAおよびヒトの進化。 の: 自然。 jg。 325、ロンドン1987、S。31–36。 PMID 3025745 doi: 10.1038/325031a0 ISSN 0028-0836
- ↑ G. David Poznik et al。: シーケンスY染色体は、男性と女性の共通の祖先との間に矛盾を解決します。 の: 化学。 バンド341、nr。 6145、2013、S。562-565、 doi:10.1126/science.1237619
- ↑ マコーレーとリチャーズ
- ↑ D.M. Behar u。 a。: Genographic Project Public Inclationation Mitochondrial DNAデータベース。 の: PLOS GENET。 jg。 3、サンフランシスコ2007、S.E104。 PMID 17604454 doi: 10.1371 / journal.pegen.0030104 ISSN 1553-7390
- ↑ R.E.緑u。 a。: ネアンデルタール語DNAの100万塩基対の分析。 の: 自然。 jg。 444、ロンドン2006、S。330–336。 PMID 17108958 doi: 10.1038/nature05336 ISSN 0028-0836
- ↑ a b 初期のヨーロッパ人はネアンデルタール人と混ざっています。 アクセス mpg.de 2015年7月12日、下顎のイラスト付き 骨1
- ↑ a b これまでに復号化された最古の現代人の環境。 2014年10月22日のMax Planck Society。
- ↑ 欧州アカデミーオブボゼン/ボルツァーノ(Eurac)、15。Januar2016 – NPO
- ↑ 遺伝子は、ネアンデルタール人からホモ・サピエンスへの流れ。 アクセス mpg.de 2015年6月28日。
- ↑ ホモサピエンスのネアンデルタルゲノム、ネアンデルタール人のサピエンスゲノムの証拠はありません。 アクセス mpg.de 2015年7月20日。
Recent Comments