Subtilis Bacillus – ウィキペディア
Subtilis Bacillus (年。 バチルス 、箸; サブティリス 、ファイン、シンプル)または Heubazillus は、広範囲にわたるグラム陽性、箸の形をした、求められている土壌細菌です。 [2] 属のすべての細菌のように バチルス は B. subtilis オプションの嫌気性内胞子。 Subtilis Bacillus 一般および応用微生物学協会によって「2023年の微生物学」が選出されました。

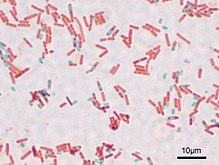
箸 B. subtilis 通常、約2〜3 µmのサイズがあり、厚さは約0.7〜0.8 µmです。その胞子は楕円形で、長さは1〜1.5 µm、幅は0.6〜0.9 µmです。 [3] 細胞はペリトリヒ、つまり数回、細胞全体に分布しているため、すぐに動くことができます。細菌はグラム陽性の細胞壁に囲まれています。 [4]
B. subtilis ユビキタスであり、地面、ほこり、空気から分離できます。土壌生殖として、植物や植物材料でしばしば発生します。 [5]
名前 Heubazillus それは、それが非常にコールされた干し草の注入で簡単に豊かになることができることを示しています。生成時間は、最適な栄養供給、最適な酸素供給、最適な成長温度40°Cで約45分です。 [4]
B. subtilis Chemoorgano Heterotroph、つまりつまり、彼は他の生物によって生成された栄養素を使用して、エネルギーと体の物質を生成します。 B. subtilis 根圏と土壌の上層の両方に定着します。 [6] そこで彼は、典型的な腐敗細菌として、食品回路に有機物質の一部を持っています。彼はグルカン(ポリマー鎖の砂糖)の大きな武器を持っています [7] とプロテア [8] 必要に応じて細胞から輸出される(タンパク質還元酵素)。
ガードシュガー(グルコース)は、炭素およびエネルギー源として好まれます。濃縮すると、グルコースは遺伝子の活性化を防ぎ、その産物は他の炭素源と代謝に干渉します。グルコース能力の場合、他の糖または炭素を含む基質も使用できます。(異化) [4]
酸素は、エネルギーを生成するための好ましい末端電子受容体(細胞呼吸)として機能します。ここでも、酸素アクセスの場合、今後の基質の使用が抑制されます。嫌気性条件下では、細胞は依然としてグルコースと硝酸の推定値の成長が遅くなるのに十分なエネルギーを生成できます。電子受容体として使用できる基質が利用可能でない場合、 B. subtilis 乳酸、エタノール、アセトイン、2.3-ブリオールの産生における発酵代謝によって排他的に生き残ることができます。 [4]

不利な環境条件は試みます B. subtilis 彼の惨劇を使用して積極的な移動を通して撤退する。さらに、できます B. subtilis 環境要因の変動を伴う栄養性活性細胞としての一般的なストレス反応について。最終的にはできます B. subtilis 非定型細胞分裂プログラムは、生態学的なニッチの放棄と進化プロセスからの逸脱にもかかわらず、長期にわたって生き残る内胞子を形成します。光顕微鏡では、胞子形成細胞の胞子または胞子は、色がなくても、強く壊れた楕円形の構造と見なすことができます。
別のプロパティは、能力のトレーニングです。バクテリアの能力は、細胞外(外来)DNAを吸収し、それを統合して独自のゲノムを拡大したり、栄養に使用したりする能力です。 [9]
分類学的にあなたを数えます B. subtilis ユーバクテリア(バクテリア)に、グラム陽性の硬化についてより正確に。そこでは、Bacilliクラス(低GC含有量)に割り当てられます。このクラスには、カリファナールと乳酸菌(乳酸酸菌)の順序が含まれます。 [十] カリファナールの順序は、以前はバチラールとして管理されていました。これには、属を持つバチラチエ科が含まれます バチルス これには、約150種が含まれます(Claus&Berkeley、1986)。ブドウ球菌とリステリア科のファミリーも、カリファナレスのオーダーの一部です。 [十] 系統系の近さは、ブドウ球菌やリステリアなどの病原性を発揮します B. subtilis 分子生物学的および医学的研究に特に興味深い。
1999年には、亜種(亜種)への分割があり、そのうち3つが現在知られています(2014年現在): [十]
- B. subtilis subsp。 inquosorum ルーニー et al。 2009年 [11] [12番目]
- B. subtilis subsp。 Spizizenii 中村 et al。 1999年 [13]
- B. subtilis subsp。 サブティリス ( エーレンバーグ 1835) 中村 et al。 1999年 [14] [13]
B. subtilis 1835年、クリスチャン・ゴットフリード・エーレンバーグとして vibrio subtilis (湾曲した箸)説明。 1872年、それはフェルディナンドジュリアスコーンによってでした Subtilis Bacillus (Stabhchen)1895年に同じ名前の家族に名前を変更して割り当てられました。 [十]
に Subtilis Bacillus グループに属します Bacillus amyloliquefaciens 、 Bacillus atrophaeus 、 Bacillus mojavensis 、 Sonorensis Bacillus 、 Bacillus tequilensis 、 Bacillus vallismortis と Bacillus velezensis 。複合体内の高度に保存されているDNA配列のため、これらは従来の16S rRNAシーケンスと区別することはできません。 [12番目] 生理学的および生化学的検査に基づく分化も不可能です。対応する特殊なデータベースがある場合、Maldi TOF MSを使用して正しい分類を実行できます。 [15]
研究で B. subtilis 細胞壁を研究するためのモデル生物として。 [16]
B. subtilis 慢性皮膚疾患(皮膚症)または下痢、発酵および腐敗性障害、胃腸炎症(腸炎)および腸炎の治療のためにヒト医学で使用されていました。ドイツで利用可能な2017年のレッドリスト(Red List、2017)では、Homeopathic Prageations UtilinとUtelin Nのみが兆候なしにリストされています。
胞子の耐熱性が高いため B. subtilis これらは、薬局、医学、食品産業における対応する滅菌プロセスの指標としても使用されます。 [17]
農業で B. subtilis STAMM QST 713生物学的農薬として商業的に使用されています。リポペプチドは、殺菌剤害虫に対して、および細菌植物の害虫に対して作用します。彼らは有機農業で承認されています。 [18]
細胞外酵素の分泌能力のため B. subtilis 特に洗剤酵素の生産のために(例:Subtilisin [19] )、ただし、リボフラビンの合成 [20] (ビタミンb 2 )および抗生物質バチトラシン [21] バイオテクノロジー産業で使用されています。
歪み Bacillus subtilis var。ナット 日本の専門のナットと同様の料理の製造に使用されます。 [22]
B. subtilis 非常にまれな状況では、病原性も機能します。 B.は、眼の損傷と細菌の浸透が発生した場合に失明につながる可能性があります(パンフタルミー)。 [23] さらに、CAN B. subtilis 毒素アミロシンを形成し、食中毒につながります。 [24]
B. subtilis 最もよく研究されたグラム陽性菌と考えられています。 1990年から1997年にかけて、彼のゲノムは研究され、完全にシーケンスされ、既存の遺伝子カードに基づいたシーケンス戦略がありました。円形のDNA二重鎖は、4,214,810塩基対で構成されています。 GC含有量は43.5%です。合計配列のうち、ヌクレオチドの86.87%がコード機能を持っています。残りのヌクレオチドはZにあります。 T.遺伝子間の調節領域。平均して、GEN 890ヌクレオチド測定。 [25]
- ↑ 種の微妙な種。 LPSN-Database、Leibniz Institute DSMZ-German微生物と細胞培養のコレクション、2023年1月3日にアクセス。
- ↑ Georg Fuchs、HansGünterSchlegel、Thomas Eitinger: 一般的な微生物学 。 8.エディション。 Thieme、Stuttgart / New York 2007、ISBN 978-3-13-44608-1、 S. 51 。
- ↑ GuntherMüller: 食物微生物学の基本:紹介 。 6日、改良版。 Steinkopf、Heidelberg 1986、ISBN 3-7985-0673-6、 S. 16 。
- ↑ a b c d Antranikian、G。(Garabed): 応用微生物学 。 Springer-Verlag Berlin Heidelberg、Berlin、Heidelberg 2006、ISBN 3-540-29456-2、 S. 14 。
- ↑ クライン、ギュンター: Microthek:Bildatlas微生物学 。 バンド 初め 。 Behr、Hamburg 2007、ISBN 978-3-89947-372-8、II.17.1。
- ↑ Shoda、Makoto: 枯草菌による植物疾患の生体上侵害:基本的および実用的なアプリケーション 。 CRC Press、Boca Raton、FL 2020、ISBN 978-0-429-63676-9、 S. 4 。
- ↑ Monika Wolf、Attila Geczi、Ortwin Simon、Rainer Borriss: バチルス亜種におけるキシランおよびβ-グルカン加水分解酵素をコードする遺伝子:リケナーゼ、セルラーゼ、キシラナーゼに不足している株の特性評価、マッピング、および構造 。の: 微生物学、 。 バンド 141 、 いいえ。 2 、1995年、ISSN 1350-0872 、 S. 281–290 、doi: 10.1099/13500872-141-2-281 。
- ↑ M. M. Kole、I。Draper、Donald F. Gerson: グルコースとアンモニウム濃度の同時制御を使用した亜種によるプロテアーゼの産生 。の: Journal of Chemical Technology&Biotechnology 。 バンド 41 、 いいえ。 3 、1988年、ISSN 1097-4660 、 S. 197–206 、doi: 10.1002/jctb.280410305 。
- ↑ PatríciaHBrito、Bastien Chevreux、Claudia R Serra、Ghislain Schyns、Adriano O Henriques、JoséBPereira-leal: 遺伝的能力は、亜種のバチルスのゲノム多様性を促進します 。の: ゲノム生物学と進化 。 バンド 十 、 いいえ。 初め 、2017、ISSN 1759-6653 、 S. 108–124 、doi: 10.1093 / place / evx270 、 PMID 29272410 。
- ↑ a b c d JeanEuzéby、Aidan C. Party: バチルスレース。 の: 命名法(LPSN)に立っている原核生物の名前のリスト。 2014年8月9日に取得 。
- ↑ Bacillus Balubed 。 doi: 10.1601/nm.14741 。
- ↑ a b Aljandro P. Rooney、Neil P. Price、Christopher Ehrhardt、James L. Swezey、Jason D. Bannan: Bacillus subtilis種の系統と分子分類法と亜ティリスbacillus subtilisの記述。 inquosorum subsp。 11月。 の: 系統的および進化的微生物学の国際ジャーナル、 。 バンド 59 、 いいえ。 十 、2009年、ISSN 1466-5026 、 S. 2429–2436 、doi: 10.1099/ijs.0.009126-0 。
- ↑ a b L. K.中村、マイケルS.ロバーツ、フレデリックM.コハン: 注:株168およびW23株に関連する亜ティリスクレードの関係:subtillus bacillus subtilis subspの提案。 Subtilis subsp。 11月。およびbacillus subtilis subsp。 Spizizenii subsp。 11月。 の: 系統的および進化的微生物学の国際ジャーナル、 。 バンド 49 、 いいえ。 3 、1999年、ISSN 1466-5026 、 S. 1211–1215 、doi: 10.1099/00207713-49-3-1211 。
- ↑ 亜種の亜種の亜種の分類法(Ehrenberg 1835)Nakamura et al。 1999年 。 doi: 10.1601/TX.4858 。
- ↑ Chien-Hsun Huang、Lina Huang、Mu-Tzu Chang、Kuo-Lung Chen: 全細胞MALDI-TOF MSテクノロジーを使用したBacillus subtilisグループ(BSG)の迅速な識別のための分析的社内データベース(IHDB)の確立と適用 。の: 分子プローブと細胞プローブ 。 バンド 30 、 いいえ。 5 、2016年、ISSN 0890-8508 、 S. 312–319 、doi: 10.1016/j.mcp.2016.08.002 。
- ↑ M. Leaver、P。Domínguez-Cuevavas、J。M。Coxhead、R。A。Daniel、J。Errington: Bacillus subtilisの壁や分割機のない生活 。の: 自然 。 バンド 457 、 いいえ。 7231 、2009年、ISSN 1476-4687 、 S. 849–853 、doi: 10.1038/nature07742 。
- ↑ T. E. Odlaug、R。A。Caputo、G。S。Graham: 凍結乾燥した亜種の胞子の耐熱性と個体群安定性 。の: 応用および環境微生物学 。 バンド 41 、 いいえ。 6 、1981年、ISSN 0099-2240 、 S. 1374–1377 、doi: 10.1128/AEM.41.6.1374-1377.1981 、 PMID 16345790 。
- ↑ ゼラー、ヴォルフガング: 細菌植物疾患の生物学的制御に関する第1回国際シンポジウムの議事録:Seehim /Darmstadt、2005年10月23日から26日 。生物学的連邦保護区。農業と林業のために、ベルリンブラウンシュヴァイグ2006、ISBN 3-930037-29-7、 S. 167 ( ResearchGate.net [PDF])。
- ↑ Sara Lanigan -Gerdes、Alek N. Dooley、Kym F. Faull、Beth A. Lazazzera: CSFを産生する酵素としてのサブチリシン、EPR、およびVPRの識別、亜種の細胞外シグナル伝達ペプチドである 。の: 分子微生物学 。 バンド 65 、 いいえ。 5 、2007年、ISSN 1365-2958 、 S. 1321–1333 、doi: 10.1111/j.1365-2958.2007.05869.x 。
- ↑ J Bパーキンス、A。スローマ、T。ヘルマン、K。セリアー、E。ザチゴ、T。エルデンバーガー、N。ハネット、N Pチャタジー、V。ウィリアムズII、GAルフォJR、R。ハッチ、J。ペロ: リボフラビンの商業生産のための亜種の遺伝子工学 。の: Journal of Industrial Microbiology and Biotechnology 。 バンド 22 、 いいえ。 初め 、1999年、ISSN 1476-5535 、 S. 8–18 、doi: 10.1038 / sj.nim.2900587 。
- ↑ Balbina A. Johnson、Herbert Anker、Frank L. Meleney: バチトラシン:B。subtilisグループのメンバーによって生成された新しい抗生物質 。の: 化学 。 バンド 102 、 いいえ。 2650 、1945年、ISSN 0036-8075 、 S. 376–377 、doi: 10.1126/science.102.2650.376 、 PMID 17770204 。
- ↑ Masahito Hitosugi, Katsuo Hamada, Kazutaka Misaka: Bacillus subtilis varの効果ライフスタイル疾患の女性患者の血流障害によって引き起こされる症状に関するnatto製品 。の: 一般医学の国際ジャーナル 。 バンド 8 、2015年、ISSN 1178-7074 、 S. 41–46 、doi: 10.2147/ijgm.s76588 、 PMID 25653551 。
- ↑ WernerKöhler(hrsg。): 医療微生物学。 8.エディション。ミュンヘン/イエナ2001、ISBN 978-3-437-41640-8。
- ↑ C. Apetroie-Constantin、R。Mikkola、M.A。 Andersson、V。Teplova、I。Suominen、T。Johansson、M。Salkkinoja-Salonen: 食中毒に接続されたBacillus subtilisとB. mojavensis株は、熱安定毒素アミロシンを生成します 。の: Journal of Applied Microbiology 。 バンド 106 、 いいえ。 6 、2009年、 S. 1976–1985 、doi: 10.1111/j.1365-2672.2009.04167.x 。
- ↑ F. Kunst、N。Ogosatrara、I。Moszer、A。Albertini、G。Alloni、V。Azevedo、M。G。Berro、P。Bessières、A。Bolotin、S。Borchet、R。Borriss、L。Boursier、M。Brans、S。Brans、S。Brans、S。Brans、S。Brans、S。 。チェ、J.-J。 Sanni、I。F. Connerton、N。J. Cummings、R。A. Daniel、F。Denizot、K。M. Devine、A。Düsterhöft、S。Ehrlich、P。Emmerson、K。D. Entian、C。Fuljitz 、、 S. Fuma、A。Galizzi、N。Galleron、S.Y。 Ghim、P。Glaser、A。Goffeau、E。J。Golightly、G。Grandi、G。Guiseppi、B。J。Guy、K。Haga、J。Haiech、C。R。Harwood、A。 Hullo、M。Heya、L。Jones、B。Joris、D。Kusara、Y。Kasahara、M。Claerr-Blanchard、C。Klein、Y。Kobayashi、P。Koetter、G。Koningstein、S。Krogh、M。K。リー、A。レヴァイン、H。リュー、S。マスダ、C。マウエル、C。メディナ、N。 Park、V。Parro、T。M。Pohl、D。Portetelle、S。Porwollik、A。Prescott、E。Presecan、P。P。Pujic、B。Purnelle、G。Rapoport、M。Rey、S。Reynolds、M。Rieger、C。Rivolta、B。Rocha、B。Rocha、 Serror、B.-S。 Shin、B。Soldo、A。Sorokin、E。Tacconi、T。Takagi、H。Takahashi、K。Takemaru、M。Tamakoshi、T。Tanaka、P。Terpstra、A。Gononi、V。Tosato、S。Vannier、F。Vassarti、A。Vassarti、A。Vassarti、A。Vasser、vasser、vassarti、A.Vassarti、A.Vassarti、A.Vassarti、 P. Winters、A。Wipat、H。Yamamoto、K。Yamane、K。Yasumoto、K。Yata、K。Yoshida、H.-F。吉川、E。ズムシュタイン、H。吉川、A。ダンチン: グラム陽性細菌菌の完全なゲノム配列 。の: 自然 。 バンド 390 、 いいえ。 6657 、1997年、ISSN 1476-4687 、 S. 249–256 、doi: 10.1038/36786 。
Recent Comments