Abscisinsäure – Wikipedia
Pflanzenhormon
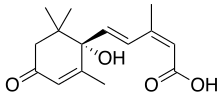 |
|
| Namen | |
|---|---|
| Bevorzugter IUPAC-Name
(2Z,4E)-5-[(1S)-1-Hydroxy-2,6,6-trimethyl-4-oxocyclohex-2-en-1-yl]-3-Methylpenta-2,4-diensäure[3]
|
|
| Andere Namen
(2Z,4E)-(S)-5-(1-Hydroxy-2,6,6-trimethyl-4-oxo-2-cyclohexen-1-yl)-3-methyl-2,4-pentandiensäure; Dorminsäure;[citation needed] Dormin[1][2]
|
|
| Bezeichner | |
| 3DMet | |
| Abkürzungen | ABA |
| 2698956 | |
| ChEBI | |
| ChEMBL | |
| ChemSpider | |
| ECHA-InfoCard | 100.040.275 |
| EG-Nummer | |
| Gittergewebe | Abscisinsäure+Säure |
| RTECS-Nummer | |
| UNII | |
|
|
|
|
| Eigenschaften | |
| Cfünfzehnh20Ö4 | |
| Molmasse | 264.321 g·mol-1 |
| Aussehen | Farblose Kristalle |
| Dichte | 1,193 g/ml |
| Schmelzpunkt | 163 °C (325 °F; 436 K)[4] |
| Protokoll P | 1.896 |
| Säure (pKein) | 4.868 |
| Basizität (pKB) | 9.129 |
| Gefahren | |
| GHS-Piktogramme |  |
| GHS-Signalwort | Warnung |
| H315, H319, H335 | |
| P261, P264, P271, P280, P302+352, P304+340, P305+351+338, P312, P321, P332+313, P337+313, P362, P403+233, P405, P501 | |
|
Sofern nicht anders angegeben, beziehen sich die Daten auf Materialien im Standardzustand (bei 25 °C [77 °F], 100 kPa).
|
|
| Infobox-Referenzen | |






Chemische Verbindung
Abscisinsäure (ABA) ist ein Pflanzenhormon. ABA funktioniert in vielen Pflanzenentwicklungsprozessen, einschließlich der Samen- und Knospenruhe, der Kontrolle der Organgröße und des Verschlusses der Stomata. Es ist besonders wichtig für Pflanzen bei der Reaktion auf Umweltstress, einschließlich Trockenheit, Bodensalzgehalt, Kältetoleranz, Gefriertoleranz, Hitzestress und Schwermetallionentoleranz.[5]
In Pflanzen[edit]
Funktion[edit]
Ursprünglich wurde angenommen, dass ABA an der Abszission beteiligt ist, weshalb es seinen Namen erhielt. Dies ist heute nur noch bei wenigen Pflanzen der Fall. ABA-vermittelte Signalübertragung spielt auch eine wichtige Rolle bei Pflanzenreaktionen auf Umweltstress und Pflanzenpathogene.[6][7] Die Pflanzengene für die ABA-Biosynthese und die Sequenz des Stoffwechselweges wurden aufgeklärt.[8][9] ABA wird auch von einigen pflanzenpathogenen Pilzen über einen anderen Biosyntheseweg als die ABA-Biosynthese in Pflanzen produziert.[10]
Zur Vorbereitung auf den Winter wird ABA in Endknospen produziert.[11] Dies verlangsamt das Pflanzenwachstum und veranlasst die Blattprimordien, Schuppen zu entwickeln, um die ruhenden Knospen während der kalten Jahreszeit zu schützen. ABA hemmt auch die Teilung von Zellen im vaskulären Kambium und passt sich an kalte Bedingungen im Winter an, indem es das primäre und sekundäre Wachstum aussetzt.
Abscisinsäure wird auch in den Wurzeln als Reaktion auf ein verringertes Bodenwasserpotential (das mit trockenem Boden verbunden ist) und andere Situationen, in denen die Pflanze unter Stress stehen kann, produziert. ABA verlagert sich dann in die Blätter, wo es das osmotische Potenzial der Stomataschließzellen schnell verändert, wodurch sie schrumpfen und sich die Stomata schließen. Der ABA-induzierte Stomataverschluss reduziert die Transpiration (Verdunstung von Wasser aus den Spaltöffnungen) und verhindert so in Zeiten geringer Wasserverfügbarkeit einen weiteren Wasserverlust aus den Blättern. Es wurde eine enge lineare Korrelation zwischen dem ABA-Gehalt der Blätter und ihrem Leitwert (Stomata-Widerstand) auf Blattflächenbasis gefunden.[12]
Die Samenkeimung wird durch ABA im Antagonismus mit Gibberellin gehemmt. ABA verhindert auch den Verlust der Samenruhe.[citation needed]
Mehrere ABA-Mutanten Arabidopsis thaliana Pflanzen wurden identifiziert und sind vom Nottingham Arabidopsis Stock Center erhältlich – sowohl solche mit einem Mangel an ABA-Produktion als auch solche mit einer veränderten Empfindlichkeit gegenüber seiner Wirkung. Pflanzen, die gegenüber ABA überempfindlich oder unempfindlich sind, zeigen Phänotypen in der Samenruhe, Keimung, Stomata-Regulation, und einige Mutanten zeigen ein verkümmertes Wachstum und braun-gelbe Blätter. Diese Mutanten spiegeln die Bedeutung von ABA bei der Samenkeimung und der frühen Embryoentwicklung wider.[citation needed]
Pyrabactin (ein Pyridyl-haltiger ABA-Aktivator) ist ein Naphthalinsulfonamid-Hypokotyl-Zellexpansionshemmer, der ein Agonist des ABA-Signalwegs der Samen ist.[13] Es ist der erste Agonist des ABA-Signalwegs, der strukturell nicht mit ABA verwandt ist.[citation needed]
Homöostase[edit]
Biosynthese[edit]
Abscisinsäure (ABA) ist ein isoprenoides Pflanzenhormon, das in der plastidalen 2-C-Methyl-D-erythritol-4-phosphat (MEP)-Weg; im Gegensatz zu den strukturell verwandten Sesquiterpenen, die aus der von Mevalonsäure abgeleiteten Vorstufe Farnesyldiphosphat (FDP) gebildet werden, ist das Cfünfzehn Rückgrat von ABA wird nach Spaltung von C . gebildet40Carotinoide in MEP. Zeaxanthin ist der erste engagierte ABA-Vorläufer; eine Reihe von enzymkatalysierten Epoxidierungen und Isomerisierungen über Violaxanthin und abschließende Spaltung des C40Carotinoid durch eine Dioxygenierungsreaktion ergibt den proximalen ABA-Vorläufer Xanthoxin, der dann weiter zu ABA oxidiert wird. über Abscisinsäurealdehyd.[8]


Abamin wurde als erster spezifischer ABA-Biosynthesehemmer entworfen, synthetisiert, entwickelt und dann patentiert, der es ermöglicht, den endogenen ABA-Spiegel zu regulieren.[14]
Ort und Zeitpunkt der ABA-Biosynthese[edit]
- Wird während der Austrocknung des vegetativen Gewebes freigesetzt und wenn Wurzeln auf Bodenverdichtung treffen.[15]
- Synthetisiert in grünen Früchten zu Beginn der Winterperiode
- In reifenden Samen synthetisiert, Ruhezustand herstellen
- Innerhalb des Blattes beweglich und kann durch den Transpirationsstrom im Xylem schnell von den Wurzeln zu den Blättern verlagert werden
- Produziert als Reaktion auf Umweltstress, wie Hitzestress, Wasserstress, Salzstress
- In allen Pflanzenteilen synthetisiert, zB Wurzeln, Blüten, Blätter und Stängel
- ABA wird in fast allen Zellen synthetisiert, die Chloroplasten oder Amyloplasten enthalten
Inaktivierung[edit]
ABA kann über CYP707A (eine Gruppe von P450-Enzymen) zu Phaseinsäure katabolisiert oder durch Glucosekonjugation (ABA-Glucoseester) über das Enzym AOG inaktiviert werden. Der Katabolismus über die CYP707As ist für die ABA-Homöostase sehr wichtig, und Mutanten in diesen Genen akkumulieren im Allgemeinen höhere ABA-Spiegel als Linien, die ABA-Biosynthesegene überexprimieren.[16] Bei Bodenbakterien wurde über einen alternativen Abbauweg berichtet, der über das Enzym Vomifoliol-Dehydrogenase zu Dehydrovomifoliol führt.
Auswirkungen[edit]
- Antitranspirant – Induziert den Verschluss der Stomata, verringert die Transpiration, um Wasserverlust zu verhindern.[17]
- Hemmt die Fruchtreife
- Verantwortlich für die Samenruhe durch Hemmung des Zellwachstums – hemmt die Samenkeimung
- Hemmt die Synthese von Kinetin-Nukleotid[18]
- Reguliert Enzyme, die für die Photosynthese benötigt werden.[19]
- Wirkt auf die Endodermis, um das Wachstum von Wurzeln zu verhindern, wenn sie salzigen Bedingungen ausgesetzt sind[20]
- Verzögert die Zellteilung
- Keimruhe-Induktor – Es wird verwendet, um die Keimruhe in den Samen zu induzieren.
- als Antitranspirant verwendet – In dürregefährdeten Gebieten ist Wasserstress ein ernstes Problem in der landwirtschaftlichen Produktion. Daher werden ABA-Sprays empfohlen, die einen teilweisen Verschluss der Stomata für einige Tage bewirken, um den Wasserverlust durch die Transpiration zu reduzieren
Signalkaskade[edit]

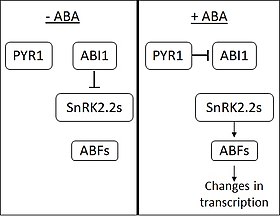
In Abwesenheit von ABA hemmt die Phosphatase ABI1-INSENSITIVE1 (ABI1) die Wirkung von SNF1-verwandten Proteinkinasen (Unterfamilie 2) (SnRK2s). ABA wird von den PYRABACTIN RESISTANCE 1 (PYR1) und PYR1-ähnlichen Membranproteinen wahrgenommen. Bei der ABA-Bindung bindet PYR1 an ABI1 und hemmt es. Wenn SnRK2s aus der Hemmung entlassen werden, aktivieren sie mehrere Transkriptionsfaktoren aus der Familie der ABA RESPONSIVE ELEMENT-BINDING FACTOR (ABF). ABFs verursachen dann Veränderungen in der Expression einer großen Anzahl von Genen.[5] Es wird angenommen, dass etwa 10 % der Pflanzengene durch ABA reguliert werden.[citation needed]
Bei Pilzen[edit]
Einige Pilzarten (z. B. Cercospora rosicola, Botrytis cinerea .)[21] und Magnaporthe oryzae) haben einen endogenen Biosyntheseweg für ABA. Bei Pilzen scheint der MVA-Biosyntheseweg vorherrschend zu sein (und nicht der MEP-Weg, der für die ABA-Biosynthese in Pflanzen verantwortlich ist). Eine Rolle der von diesen Pathogenen produzierten ABA scheint darin zu bestehen, die pflanzlichen Immunantworten zu unterdrücken.[22]
Bei Tieren[edit]
ABA wurde auch in Metazoen gefunden, von Schwämmen bis hin zu Säugetieren einschließlich des Menschen.[23] Derzeit ist seine Biosynthese und biologische Rolle bei Tieren wenig bekannt. Kürzlich wurde gezeigt, dass ABA starke entzündungshemmende und antidiabetische Wirkungen in Mausmodellen für Diabetes/Fettleibigkeit, entzündliche Darmerkrankungen, Arteriosklerose und Grippeinfektionen hervorruft.[24] Viele biologische Wirkungen bei Tieren wurden unter Verwendung von ABA als nutrazeutisches oder pharmakognostisches Medikament untersucht, aber ABA wird auch von einigen Zellen (wie Makrophagen) endogen erzeugt, wenn sie stimuliert werden. Es gibt auch widersprüchliche Schlussfolgerungen aus verschiedenen Studien, in denen einige behaupten, dass ABA für entzündungsfördernde Reaktionen unerlässlich ist, während andere entzündungshemmende Wirkungen zeigen. Wie viele Naturstoffe mit medizinischen Eigenschaften hat sich ABA auch in der Naturheilkunde durchgesetzt. Während ABA eindeutig positive biologische Aktivitäten hat und viele naturheilkundliche Mittel einen hohen Gehalt an ABA enthalten (wie Weizengrassaft, Obst und Gemüse), können einige der gesundheitsbezogenen Angaben übertrieben oder zu optimistisch sein. In Säugerzellen zielt ABA auf ein Protein, das als Lanthioninsynthetase C-like 2 (LANCL2) bekannt ist, und löst einen alternativen Mechanismus der Aktivierung des Peroxisom-Proliferator-aktivierten Gamma-Rezeptors (PPAR-Gamma) aus.[25] LANCL2 ist in Pflanzen konserviert und wurde ursprünglich auch in Pflanzen als ABA-Rezeptor vorgeschlagen, was später herausgefordert wurde.[26]
Messung der ABA-Konzentration[edit]
Mehrere Methoden können helfen, die Konzentration von Abscisinsäure in einer Vielzahl von Pflanzengeweben zu quantifizieren. Die verwendeten quantitativen Methoden basieren auf HPLC und GC sowie ELISA. Vor kurzem wurden zwei unabhängige FRET-Sonden entwickelt, die intrazelluläre ABA-Konzentrationen in Echtzeit in vivo messen können.[27][28]
Verweise[edit]
- ^ O’Neil, Maryadele J; Heckelmann, PE; Koch, CB; Roman, KJ (2006). Der Merck-Index, 14..
- ^ 21293-29-8
- ^ “Abscisinsäure – Zusammenfassung der Verbindungen”. PubChem-Verbindung. USA: Nationales Zentrum für Biotechnologie-Informationen. 16. September 2004. Identifizierung und zugehörige Aufzeichnungen. Abgerufen 22. Oktober 2011.
- ^ “ChemSpider-Datenbank – Abscisinsäure – Eigenschaften”. Abgerufen 27. Dezember 2012. Der Schmelzpunkt wird durch experimentelle Daten von Tokyo Chemical Industry Ltd. bestimmt.
- ^ ein B Finkelstein, Ruth (2013-11-01). “Abscisinsäure Synthese und Reaktion”. Das Arabidopsis-Buch / American Society of Plant Biologists. 11: e0166. mach:10.1199/tab.0166. PMC 3833200. PMID 24273463.
- ^ Zhu, Jian-Kang (2002). “Salz- und Trockenstress-Signaltransduktion in Pflanzen”. Jährliche Überprüfung der Pflanzenbiologie. 53: 247–73. mach:10.1146/annurev.arplant.53.091401.143329. PMC 3128348. PMID 12221975.
- ^ SEO, M; Koshiba, T (2002). „Komplexe Regulierung der ABA-Biosynthese in Pflanzen“. Trends in der Pflanzenwissenschaft. 7 (1): 41–8. mach:10.1016/S1360-1385(01)02187-2. PMID 11804826.
- ^ ein B Nambara, Eiji; Marion-Poll, Annie (2005). „Abscisinsäure Biosynthese und Katabolismus“. Jährliche Überprüfung der Pflanzenbiologie. 56: 165–85. mach:10.1146/annurev.arplant.56.032604.144046. PMID 15862093.
- ^ Milborrow, BV (2001). “Der Weg der Biosynthese von Abscisinsäure in Gefäßpflanzen: Ein Überblick über den aktuellen Wissensstand der ABA-Biosynthese”. Zeitschrift für experimentelle Botanik. 52 (359): 1145–64. mach:10.1093/jexbot/52.359.1145. PMID 11432933.
- ^
- ^ Wang, Dongling; Gao, Zhenzen; Du, Peiyong; Xiao, Wei; Bräune, Qiuping; Chen, Xiude; Li, Ling; Gao, Dongsheng (2016). “Expression von ABA-Stoffwechsel-bezogenen Genen deutet auf Ähnlichkeiten und Unterschiede zwischen der Samenruhe und der Knospenruhe von Pfirsich (Prunus persica) hin”. Grenzen in der Pflanzenwissenschaft. 6: 1248. doi:10.3389/fpls.2015.01248. ISSN 1664-462X. PMC 4707674. PMID 26793222.
- ^ Steuer, Barbara; Thomas Stuhlfauth; Heinrich P. Fock (1988). “Die Effizienz der Wassernutzung in wassergestressten Pflanzen wird durch den ABA-induzierten Stomataverschluss erhöht”. Photosyntheseforschung. 18 (3): 327–336. mach:10.1007/BF00034837. ISSN 0166-8595. PMID 24425243. S2CID 30298332.[citation needed]
- ^ Park, Sang-Youl; P. Fung; N. Nishimura; DR. Jensen; H. Fuji; Y. Zhao, S. Lumba; et al. (Mai 2009). „Abscisinsäure hemmt Proteinphosphatasen vom Typ 2C über die PYR/PYL-Familie der START-Proteine“. Wissenschaftliche Signalgebung. 324 (5930): 1068–1071. Bibcode:2009Sci…324.1068P. mach:10.1126/science.1173041. PMC 2827199. PMID 19407142.
- ^ Abscisinsäure-Biosynthesehemmer, Shigeo Yoshida et al US 7098365
- ^ DeJong-Hughes, J., et al. (2001)Bodenverdichtung: Ursachen, Wirkungen und Kontrolle. Erweiterungsservice der University of Minnesota
- ^ Finkelstein, Ruth (November 2013). “Abscisinsäure Synthese und Reaktion”. Das Arabidopsis-Buch. 11: e0166. mach:10.1199/tab.0166. PMC 3833200. PMID 24273463.
- ^ Zhang, Jianhua; Schurr, U.; Davies, WJ (1987). „Kontrolle des Stomatalverhaltens durch Abscisinsäure, die anscheinend in den Wurzeln entsteht“. Zeitschrift für experimentelle Botanik. 38 (7): 1174–1181. mach:10.1093/jxb/38.7.1174.
- ^ Miernyk, JA (1979). „Abscisinsäure Hemmung der Kinetin-Nukleotid-Bildung in keimenden Salatsamen“. Physiologia Plantarum. 45: 63–6. mach:10.1111/j.1399-3054.1979.tb01664.x.
- ^ Krämer, PM; Robertson, M. (1994). „Genexpression durch Abscisinsäure und ihre Beziehung zur Stresstoleranz reguliert“. Jährlicher Überblick über Pflanzenphysiologie und Pflanzenmolekularbiologie. 45: 113–41. mach:10.1146/annurev.pp.45.060194.000553.
- ^ Duan, Lina; D. Dietrich; CHNg; PMY-Chan; R. Bhalerao; MJ Bennett; JR Dinneny. (Januar 2013). “Endodermale ABA-Signalgebung fördert die seitliche Wurzelruhe bei Salzstress bei Arabidopsis-Sämlingen”. Die Pflanzenzelle. 25 (1): 324–341. mach:10.1105/tSt.112.107227. PMC 3584545. PMID 23341337.
- ^ Sievers, Verena; Kokkelink, Leonie; Smedsgaard, Jørn; Tudzynski, Paul (Juli 2006). “Identifizierung eines Abscisinsäure-Genclusters im Grauschimmel Botrytis cinerea”. Appl Environ Microbiol. 72 (7): 4619–4626. Bibcode:2006ApEnM..72.4619S. mach:10.1128/AEM.02919-05. PMC 1489360. PMID 16820452.
- ^ Lievens, Laurens; Pollier, Jakob; Goossens, Alain; Beyaert, Rudi; Staal, Jens (2017). „Abscisinsäure als Krankheitserreger und Immunregulator“. Grenzen in der Pflanzenwissenschaft. 8: 587. doi:10.3389/fpls.2017.00587. ISSN 1664-462X. PMC 5395610. PMID 28469630.
- ^ Na-Hang, Li; Rui-Lin, Hao; Shan-Shan, Wu; Peng-Cheng, Guo; Can-Jiang, Chen; Li-Ping, Pan; Er, Ni (2011). „Vorkommen, Funktion und mögliche medizinische Anwendungen des Phytohormons Abscisinsäure bei Tieren und Menschen“. Biochemische Pharmakologie. 82 (7): 701–712. mach:10.1016/j.bcp.2011.06.042. PMID 21763293.
- ^ Bassaganya-Riera, J; Skoneczka, J; Kingston, DG; Krishnan, A; Misyak, SA; Guri, AJ; Pereira, A; Carter, AB; Minorski, P; Tumarkin, R; Hontecillas, R (2010). “Wirkmechanismen und medizinische Anwendungen von Abscisinsäure”. Aktuelle medizinische Chemie. 17 (5): 467–78. mach:10.2174/092986710790226110. PMID 20015036. Archiviert von das Original am 2012-04-01. Abgerufen 2018-09-30.
- ^ Bassaganya-Riera, J.; Guri, AJ; Lu, P.; Climent, M.; Carbo, A.; Sobral, BW; Horne, WT; Lewis, SN; Bevan, DR; Hontecillas, R. (2010). „Abscisinsäure reguliert Entzündungen über Liganden-bindende Domänen-unabhängige Aktivierung des Peroxisom-Proliferator-aktivierten Rezeptors“. Zeitschrift für biologische Chemie. 286 (4): 2504–16. mach:10.1074/jbc.M110.160077. PMC 3024745. PMID 21088297.
- ^ Chen, JG; Ellis, BE (2008). „GCR2 ist ein neues Mitglied der eukaryontischen Lanthioninsynthetase-Komponenten-C-like-Proteinfamilie“. Signalverhalten von Pflanzen. 3 (5): 307–10. mach:10.4161/psb.3.5.5292. PMC 2634266. PMID 19841654.
- ^ Waadt, R; Hitomi, K; Nishimura, N; Hitomi, C; Adams, SR; Getzoff, ED; Schröder, JI (2014). “FRET-basierte Reporter zur direkten Visualisierung von Konzentrationsänderungen und Verteilung von Abscisinsäure in Arabidopsis”. eLife. 3: e01739. mach:10.7554/eLife.01739. PMC 3985518. PMID 24737861.
- ^ Jones, AM; Danielson, JA; Manjokumar, SN; Laquar, V.; Großmann, G; Frommer, WB (2014). “Abscisinsäure-Dynamik in Wurzeln mit genetisch kodierten FRET-Sensoren nachgewiesen”. eLife. 3: e01741. mach:10.7554/eLife.01741. PMC 3985517. PMID 24737862.
Recent Comments