Nahrungsnetz – Wikipedia
Natürliche Verbindung von Nahrungsketten
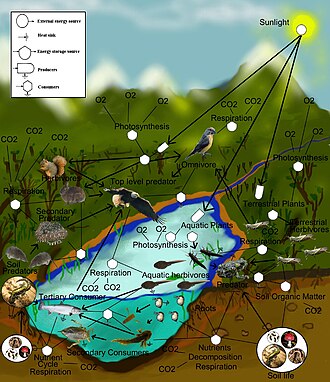
EIN Nahrungsnetz (oder Lebensmittelkreislauf) ist die natürliche Verbindung von Nahrungsketten und eine grafische Darstellung (normalerweise ein Bild) dessen, was in einer ökologischen Gemeinschaft was isst. Ein anderer Name für Nahrungsnetz ist Verbraucher-Ressourcensystem. Ökologen können alle Lebensformen grob in eine von zwei Kategorien einteilen, die als trophische Ebenen bezeichnet werden: 1) die Autotrophen und 2) die Heterotrophen. Um ihren Körper zu erhalten, zu wachsen, sich zu entwickeln und sich zu vermehren, produzieren Autotrophe organische Stoffe aus anorganischen Substanzen, einschließlich Mineralien und Gasen wie Kohlendioxid. Diese chemischen Reaktionen erfordern Energie, die hauptsächlich von der Sonne und größtenteils durch Photosynthese stammt, obwohl eine sehr geringe Menge aus der Bioelektrogenese in Feuchtgebieten stammt.[1] und Mineralelektronendonoren in hydrothermalen Quellen und heißen Quellen. Es besteht ein Gradient zwischen trophischen Konzentrationen, die von vollständigen Autotrophen reichen, die ihre einzige Kohlenstoffquelle aus der Atmosphäre beziehen, bis zu Mixotrophen (wie fleischfressenden Pflanzen), die autotrophe Organismen sind, die teilweise organische Stoffe aus anderen Quellen als der Atmosphäre erhalten, und vollständigen Heterotrophen, die dies müssen füttern, um organische Substanz zu erhalten. Die Verknüpfungen in einem Nahrungsnetz veranschaulichen die Fütterungswege, beispielsweise wo Heterotrophe organische Substanz erhalten, indem sie sich von Autotrophen und anderen Heterotrophen ernähren. Das Nahrungsnetz ist eine vereinfachte Darstellung der verschiedenen Fütterungsmethoden, die ein Ökosystem zu einem einheitlichen Austauschsystem verbinden. Es gibt verschiedene Arten von Fütterungsbeziehungen, die grob in Pflanzenfresser, Fleischfresser, Aasfresser und Parasitismus unterteilt werden können. Ein Teil der organischen Substanz, die von Heterotrophen wie Zucker aufgenommen wird, liefert Energie. Autotrophe und Heterotrophe kommen in allen Größen vor, von mikroskopisch bis zu vielen Tonnen – von Cyanobakterien bis zu Riesenmammutbäumen und von Viren und Bdellovibrio bis zu Blauwalen.
Charles Elton war in seinem klassischen Buch “Animal Ecology” von 1927 Pionier des Konzepts von Nahrungszyklen, Nahrungsketten und Nahrungsgrößen. Eltons “Nahrungszyklus” wurde in einem nachfolgenden ökologischen Text durch “Nahrungsnetz” ersetzt. Elton organisierte Arten in funktionelle Gruppen, was die Grundlage für Raymond Lindemans klassisches und wegweisendes Papier über trophische Dynamik im Jahr 1942 war. Lindeman betonte die wichtige Rolle von Zersetzerorganismen in einem trophischen Klassifikationssystem. Der Begriff eines Nahrungsnetzes hat einen historischen Halt in den Schriften von Charles Darwin und seiner Terminologie, einschließlich einer “verschränkten Bank”, eines “Netzes des Lebens”, eines “Netzes komplexer Beziehungen” und in Bezug auf die Zersetzungsaktionen von Regenwürmern, die er hat sprach über “die fortgesetzte Bewegung der Teilchen der Erde”. Noch früher, 1768, beschrieb John Bruckner die Natur als “ein fortbestehendes Netz des Lebens”.
Nahrungsnetze sind begrenzte Darstellungen realer Ökosysteme, da sie notwendigerweise viele Arten zu trophischen Arten zusammenfassen, die funktionelle Gruppen von Arten sind, die dieselben Raubtiere und Beute in einem Nahrungsnetz haben. Ökologen verwenden diese Vereinfachungen in quantitativen (oder mathematischen) Modellen der Dynamik trophischer oder Verbraucherressourcensysteme. Mit diesen Modellen können sie verallgemeinerte Muster in der Struktur realer Nahrungsnetzwerke messen und testen. Ökologen haben nicht zufällige Eigenschaften in der topografischen Struktur von Nahrungsnetzen identifiziert. Veröffentlichte Beispiele, die in der Metaanalyse verwendet werden, sind von variabler Qualität mit Auslassungen. Die Zahl der empirischen Studien zu Community-Netzen nimmt jedoch zu, und die mathematische Behandlung von Nahrungsnetzen mithilfe der Netzwerktheorie hat Muster identifiziert, die allen gemeinsam sind. Skalierungsgesetze sagen beispielsweise eine Beziehung zwischen der Topologie der Verbindungen zwischen Raubtier und Beute im Nahrungsnetz und dem Grad des Artenreichtums voraus.
Taxonomie eines Nahrungsnetzes[edit]

Links in Nahrungsnetzen bilden die Fütterungsverbindungen (wer isst wen) in einer ökologischen Gemeinschaft ab. Lebensmittelkreislauf ist ein veralteter Begriff, der gleichbedeutend mit Nahrungsnetz ist. Ökologen können alle Lebensformen grob in eine von zwei trophischen Schichten einteilen, die Autotrophen und die Heterotrophen. Autotrophe produzieren mehr Biomasseenergie, entweder chemisch ohne Sonnenenergie oder durch Erfassung der Sonnenenergie bei der Photosynthese, als sie während der Stoffwechselatmung verbrauchen. Heterotrophe verbrauchen eher Biomasseenergie als diese zu produzieren, da sie metabolisieren, wachsen und die Sekundärproduktion steigern. Ein Nahrungsnetz zeigt eine Sammlung polyphager heterotropher Verbraucher, die den Energie- und Nährstofffluss aus einer produktiven Basis selbsternährender Autotrophen vernetzen und zyklisieren.[4][5][6]
Die Basis- oder Basalarten in einem Nahrungsnetz sind solche Arten ohne Beute und können Autotrophen oder saprophytische Detritivoren umfassen (dh die Gemeinschaft von Zersetzern in Boden, Biofilmen und Periphyton). Feeding-Verbindungen im Web werden als trophische Links bezeichnet. Die Anzahl der trophischen Verbindungen pro Verbraucher ist ein Maß für die Verbindung des Nahrungsnetzes. Nahrungsketten sind in den trophischen Gliedern von Nahrungsnetzen verschachtelt. Nahrungsketten sind lineare (nichtzyklische) Nahrungswege, die monophagische Konsumenten von einer Basisspezies bis zum Topkonsumenten verfolgen, der normalerweise ein größerer räuberischer Fleischfresser ist.[7][8][9]
Verknüpfungen verbinden sich mit Knoten in einem Nahrungsnetz, bei denen es sich um Aggregate biologischer Taxa handelt, die als trophische Arten bezeichnet werden. Trophäenarten sind funktionelle Gruppen, die dieselben Raubtiere und Beute in einem Nahrungsnetz haben. Häufige Beispiele für einen aggregierten Knoten in einem Nahrungsnetz können Parasiten, Mikroben, Zersetzer, Saprotrophen, Verbraucher oder Raubtiere sein, die jeweils viele Arten in einem Netz enthalten, die ansonsten mit anderen trophischen Arten verbunden werden können.[10][11]
Trophäenlevel[edit]
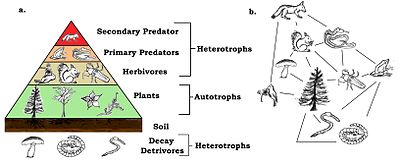
Nahrungsnetze haben trophische Ebenen und Positionen. Basalarten wie Pflanzen bilden die erste Ebene und sind die ressourcenbeschränkten Arten, die sich von keinem anderen Lebewesen im Netz ernähren. Basalspezies können Autotrophen oder Detritivoren sein, einschließlich “Zersetzung von organischem Material und den damit verbundenen Mikroorganismen, die wir als Detritus, mikroanorganisches Material und assoziierte Mikroorganismen (MIP) sowie Gefäßpflanzenmaterial definiert haben”.[12]::94 Die meisten Autotrophen fangen die Sonnenenergie in Chlorophyll ein, aber einige Autotrophen (die Chemolithotrophen) gewinnen Energie durch chemische Oxidation anorganischer Verbindungen und können in dunklen Umgebungen wie dem Schwefelbakterium wachsen Thiobacillus, die in heißen Schwefelquellen lebt. Die oberste Ebene hat Top- (oder Apex-) Raubtiere, die keine andere Art direkt für ihren Nahrungsbedarf tötet. Die Zwischenstufen sind mit Allesfressern gefüllt, die sich von mehr als einer trophischen Stufe ernähren und dafür sorgen, dass Energie ausgehend von einer Basalart durch eine Reihe von Nahrungswegen fließt.[13]
Im einfachsten Schema besteht die erste trophische Stufe (Stufe 1) aus Pflanzen, dann aus Pflanzenfressern (Stufe 2) und dann aus Fleischfressern (Stufe 3). Die trophische Ebene ist gleich eins mehr als die Kettenlänge, dh die Anzahl der Glieder, die mit der Basis verbunden sind. Die Basis der Nahrungskette (Primärproduzenten oder Detritivoren) wird auf Null gesetzt.[4][14] Ökologen identifizieren Fütterungsbeziehungen und organisieren Arten in trophische Arten durch umfassende Analyse des Darminhalts verschiedener Arten. Die Technik wurde durch die Verwendung stabiler Isotope verbessert, um den Energiefluss durch die Bahn besser verfolgen zu können.[15] Es wurde einmal angenommen, dass Allesfresser selten sind, aber neuere Beweise deuten auf etwas anderes hin. Diese Erkenntnis hat die trophischen Klassifikationen komplexer gemacht.[16]
Trophäendynamik[edit]
Das Konzept der trophischen Ebene wurde 1942 von Raymond L. Lindeman in einem historischen Meilensteinpapier zur trophischen Dynamik vorgestellt. Die Grundlage der trophischen Dynamik ist die Übertragung von Energie von einem Teil des Ökosystems auf einen anderen.[14][17] Das trophische dynamische Konzept hat als nützliche quantitative Heuristik gedient, weist jedoch einige wesentliche Einschränkungen auf, einschließlich der Genauigkeit, mit der ein Organismus einer bestimmten trophischen Ebene zugeordnet werden kann. Allesfresser zum Beispiel sind nicht auf eine einzelne Ebene beschränkt. Neuere Forschungen haben jedoch herausgefunden, dass es diskrete trophische Ebenen gibt, aber “oberhalb der trophischen Ebene von Pflanzenfressern werden Nahrungsnetze besser als ein Wirrwarr von Allesfressern charakterisiert.”[16]
Eine zentrale Frage in der trophischen dynamischen Literatur ist die Art der Kontrolle und Regulierung von Ressourcen und Produktion. Ökologen verwenden vereinfachte Modelle der Nahrungskette mit einer trophischen Position (Produzent, Fleischfresser, Zersetzer). Mit diesen Modellen haben Ökologen verschiedene Arten von ökologischen Kontrollmechanismen getestet. Zum Beispiel haben Pflanzenfresser im Allgemeinen eine Fülle von vegetativen Ressourcen, was bedeutete, dass ihre Populationen weitgehend von Raubtieren kontrolliert oder reguliert wurden. Dies ist als Top-Down-Hypothese oder “Green-World” -Hypothese bekannt. Alternativ zur Top-Down-Hypothese ist nicht das gesamte Pflanzenmaterial essbar, und die Nährstoffqualität oder die Abwehr gegen Pflanzenfresser von Pflanzen (strukturell und chemisch) deuten auf eine Bottom-up-Form der Regulierung oder Kontrolle hin.[18][19][20] Jüngste Studien haben ergeben, dass sowohl “Top-Down” – als auch “Bottom-Up” -Kräfte die Gemeinschaftsstruktur beeinflussen können und die Stärke des Einflusses vom Kontext abhängt.[21][22] Diese komplexen multitrophen Wechselwirkungen betreffen mehr als zwei trophische Ebenen in einem Nahrungsnetz.[23]
Ein weiteres Beispiel für eine multitrophische Interaktion ist eine trophische Kaskade, bei der Raubtiere durch Unterdrückung von Pflanzenfressern das Pflanzenwachstum steigern und eine Überweidung verhindern. Links in einem Nahrungsnetz veranschaulichen direkte trophische Beziehungen zwischen Arten, aber es gibt auch indirekte Effekte, die die Häufigkeit, Verteilung oder Biomasse in den trophischen Ebenen verändern können. Zum Beispiel beeinflussen Raubtiere, die Pflanzenfresser essen, indirekt die Kontrolle und Regulierung der Primärproduktion in Pflanzen. Obwohl die Raubtiere die Pflanzen nicht direkt fressen, regulieren sie die Population von Pflanzenfressern, die direkt mit dem Pflanzentrophäismus verbunden sind. Der Nettoeffekt direkter und indirekter Beziehungen wird als trophische Kaskaden bezeichnet. Trophäenkaskaden werden in Kaskaden auf Artenebene unterteilt, bei denen nur eine Teilmenge der Dynamik des Nahrungsnetzes von einer Änderung der Bevölkerungszahlen betroffen ist, und Kaskaden auf Gemeindeebene, bei denen eine Änderung der Bevölkerungszahlen dramatische Auswirkungen auf die gesamte Lebensmittelzahl hat. Web, wie die Verteilung der pflanzlichen Biomasse.[24]
Energiefluss und Biomasse[edit]
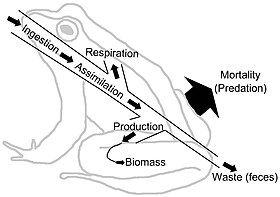
Nahrungsnetze zeigen den Energiefluss über trophische Verknüpfungen. Der Energiefluss ist gerichtet, was im Gegensatz zu den zyklischen Materialflüssen durch die Nahrungsnetzsysteme steht.[27] Der Energiefluss “umfasst typischerweise Produktion, Verbrauch, Assimilation, Nichtassimilationsverluste (Kot) und Atmung (Wartungskosten).”[6]::5 In einem sehr allgemeinen Sinne kann der Energiefluss (E) als die Summe aus Stoffwechselproduktion (P) und Atmung (R) definiert werden, so dass E = P + R.
Biomasse steht für gespeicherte Energie. Die Konzentration und Qualität von Nährstoffen und Energie ist jedoch unterschiedlich. Beispielsweise sind viele Pflanzenfasern für viele Pflanzenfresser unverdaulich, so dass Nahrungsnetze in der Grazer-Gemeinschaft nährstoffbeschränkter sind als schädliche Nahrungsnetze, in denen Bakterien auf die Nährstoff- und Energiespeicher zugreifen und diese freisetzen können.[28][29] “Organismen extrahieren normalerweise Energie in Form von Kohlenhydraten, Lipiden und Proteinen. Diese Polymere spielen eine doppelte Rolle als Energieversorgung sowie als Bausteine. Der Teil, der als Energieversorgung fungiert, führt zur Produktion von Nährstoffen (und Kohlendioxid). Wasser und Wärme). Die Ausscheidung von Nährstoffen ist daher für den Stoffwechsel von grundlegender Bedeutung. “[29]::1230–1231 Die Einheiten in Energieflussnetzen sind typischerweise ein Maß für Masse oder Energie pro m2 pro Zeiteinheit. Unterschiedliche Verbraucher werden unterschiedliche Stoffwechselassimilationseffizienzen in ihrer Ernährung haben. Jede trophische Ebene wandelt Energie in Biomasse um. Energieflussdiagramme veranschaulichen die Geschwindigkeit und Effizienz des Transfers von einer trophischen Ebene in eine andere und durch die Hierarchie.[30][31]
Es ist der Fall, dass die Biomasse jeder trophischen Ebene von der Basis der Kette nach oben abnimmt. Dies liegt daran, dass bei jeder Übertragung mit zunehmender Entropie Energie an die Umwelt verloren geht. Etwa achtzig bis neunzig Prozent der Energie werden für die Lebensprozesse des Organismus aufgewendet oder gehen als Wärme oder Abfall verloren. Nur etwa zehn bis zwanzig Prozent der Energie des Organismus werden im Allgemeinen an den nächsten Organismus weitergegeben.[32] Die Menge kann bei Tieren, die weniger verdauliche Pflanzen konsumieren, weniger als ein Prozent betragen, und bei Zooplankton, das Phytoplankton konsumiert, kann sie bis zu vierzig Prozent betragen.[33] Grafische Darstellungen der Biomasse oder Produktivität auf jeder Tropenebene werden als ökologische Pyramiden oder trophische Pyramiden bezeichnet. Der Energietransfer von Primärproduzenten zu Top-Verbrauchern kann auch durch Energieflussdiagramme charakterisiert werden.[34]
Nahrungskette[edit]
Eine übliche Metrik zur Quantifizierung der trophischen Struktur des Nahrungsnetzes ist die Länge der Nahrungskette. Die Länge der Nahrungskette ist eine weitere Möglichkeit, Nahrungsnetze als Maß für die Anzahl der Arten zu beschreiben, die auftreten, wenn Energie oder Nährstoffe von den Pflanzen zu den Top-Raubtieren gelangen.[35]::269 Es gibt verschiedene Methoden zur Berechnung der Länge der Nahrungskette, je nachdem, welche Parameter der Dynamik des Nahrungsnetzes berücksichtigt werden: Konnektivität, Energie oder Interaktion.[35] In seiner einfachsten Form ist die Länge einer Kette die Anzahl der Glieder zwischen einem trophischen Verbraucher und der Basis des Webs. Die mittlere Kettenlänge eines gesamten Netzes ist der arithmetische Durchschnitt der Längen aller Ketten in einem Nahrungsnetz.[36][13]
In einem einfachen Raubtier-Beute-Beispiel ist ein Hirsch einen Schritt von den Pflanzen entfernt, die er frisst (Kettenlänge = 1), und ein Wolf, der den Hirsch frisst, ist zwei Schritte von den Pflanzen entfernt (Kettenlänge = 2). Die relative Menge oder Stärke des Einflusses, den diese Parameter auf das Nahrungsnetz haben, beantwortet Fragen zu:
- die Identität oder Existenz einiger weniger dominanter Arten (sogenannte starke Interaktoren oder Keystone-Arten)
- die Gesamtzahl der Arten und die Länge der Nahrungskette (einschließlich vieler schwacher Interaktoren) und
- wie die Struktur, Funktion und Stabilität der Gemeinschaft bestimmt wird.[37][38]
Ökologische Pyramiden[edit]
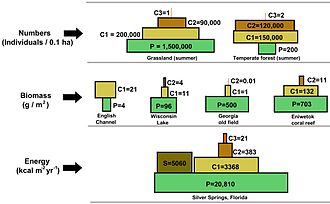
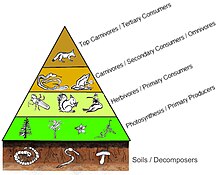
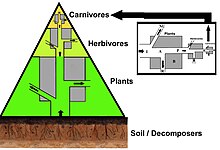
In einer Pyramide von Zahlen nimmt die Anzahl der Verbraucher auf jeder Ebene erheblich ab, so dass ein einzelner Top-Verbraucher (z. B. ein Eisbär oder ein Mensch) von einer viel größeren Anzahl separater Hersteller unterstützt wird. In einer Nahrungskette gibt es normalerweise maximal vier oder fünf Glieder, obwohl die Nahrungsketten in aquatischen Ökosystemen häufiger länger sind als an Land. Schließlich wird die gesamte Energie in einer Nahrungskette als Wärme verteilt.[5]
Ökologische Pyramiden stellen die Primärproduzenten an die Basis. Sie können verschiedene numerische Eigenschaften von Ökosystemen darstellen, einschließlich der Anzahl der Individuen pro Flächeneinheit, Biomasse (g / m)2) und Energie (k cal m−2 Jahr−1). Die sich abzeichnende pyramidenförmige Anordnung der trophischen Ebenen mit abnehmender Energieübertragung, wenn Arten weiter von der Produktionsquelle entfernt werden, ist eines von mehreren Mustern, die sich in den Ökosystemen der Planeten wiederholen.[3][4][39] Die Größe jeder Ebene in der Pyramide repräsentiert im Allgemeinen Biomasse, die als Trockengewicht eines Organismus gemessen werden kann.[40] Autotrophe haben zwar den weltweit höchsten Anteil an Biomasse, werden jedoch von Mikroben stark konkurriert oder übertroffen.[41][42]
Die Pyramidenstruktur kann je nach Ökosystem und Zeit variieren. In einigen Fällen können Biomassepyramiden invertiert werden. Dieses Muster wird häufig in aquatischen und Korallenriff-Ökosystemen identifiziert. Das Muster der Inversion von Biomasse wird auf unterschiedliche Größen von Erzeugern zurückgeführt. Wassergemeinschaften werden häufig von Produzenten dominiert, die kleiner sind als die Verbraucher mit hohen Wachstumsraten. Wasserproduzenten wie Planktonalgen oder Wasserpflanzen fehlt die große Anhäufung von Sekundärwachstum, wie sie in den Holzbäumen terrestrischer Ökosysteme vorhanden ist. Sie können sich jedoch schnell genug vermehren, um eine größere Biomasse von Weiden zu unterstützen. Dies kehrt die Pyramide um. Primärverbraucher haben eine längere Lebensdauer und langsamere Wachstumsraten, die mehr Biomasse ansammeln als die von ihnen konsumierten Produzenten. Phytoplankton lebt nur wenige Tage, während das Zooplankton, das das Phytoplankton frisst, mehrere Wochen lebt und die Fische, die das Zooplankton fressen, mehrere aufeinanderfolgende Jahre leben.[43] Aquatische Raubtiere haben tendenziell auch eine niedrigere Sterblichkeitsrate als die kleineren Verbraucher, was zum umgekehrten Pyramidenmuster beiträgt. Bevölkerungsstruktur, Migrationsraten und Umweltschutz für Beute sind weitere mögliche Ursachen für Pyramiden mit umgekehrter Biomasse. Energiepyramiden haben jedoch immer eine aufrechte Pyramidenform, wenn alle Energiequellen für Lebensmittel enthalten sind, und dies wird durch den zweiten Hauptsatz der Thermodynamik vorgegeben.[5][44]
Materialfluss und Recycling[edit]
Viele der Elemente und Mineralien der Erde (oder Mineralstoffe) sind im Gewebe und in der Ernährung von Organismen enthalten. Daher verfolgen Mineral- und Nährstoffkreisläufe die Energiepfade des Nahrungsnetzes. Ökologen verwenden Stöchiometrie, um die Verhältnisse der Hauptelemente aller Organismen zu analysieren: Kohlenstoff (C), Stickstoff (N), Phosphor (P). Es gibt einen großen Übergangsunterschied zwischen vielen terrestrischen und aquatischen Systemen, da die C: P- und C: N-Verhältnisse in terrestrischen Systemen viel höher sind, während die N: P-Verhältnisse zwischen den beiden Systemen gleich sind.[45][46][47]Mineralische Nährstoffe sind die materiellen Ressourcen, die Organismen für Wachstum, Entwicklung und Vitalität benötigen. Nahrungsnetze zeigen die Wege des Mineralstoffkreislaufs, während sie durch Organismen fließen.[5][17] Der größte Teil der Primärproduktion in einem Ökosystem wird nicht verbraucht, sondern durch Abfall wieder in nützliche Nährstoffe umgewandelt.[48] Viele der Mikroorganismen der Erde sind an der Bildung von Mineralien in einem Prozess beteiligt, der als Biomineralisierung bezeichnet wird.[49][50][51] Bakterien, die in schädlichen Sedimenten leben, erzeugen und zirkulieren Nährstoffe und Biomineralien.[52] Nahrungsnetzmodelle und Nährstoffkreisläufe wurden traditionell getrennt behandelt, es besteht jedoch eine starke funktionale Verbindung zwischen beiden in Bezug auf Stabilität, Flussmittel, Quellen, Senken und Recycling von Mineralstoffen.[53][54]
Arten von Nahrungsnetzen[edit]
Nahrungsnetze sind notwendigerweise aggregiert und veranschaulichen nur einen winzigen Teil der Komplexität realer Ökosysteme. Zum Beispiel liegt die Anzahl der Arten auf dem Planeten wahrscheinlich in der allgemeinen Größenordnung von 107Über 95% dieser Arten bestehen aus Mikroben und Wirbellosen, und relativ wenige wurden von Taxonomen benannt oder klassifiziert.[55][56][57] Es wird ausdrücklich verstanden, dass natürliche Systeme „schlampig“ sind und dass trophische Positionen im Nahrungsnetz die Komplexität realer Systeme vereinfachen, die manchmal viele seltene Wechselwirkungen überbetonen. Die meisten Studien konzentrieren sich auf die größeren Einflüsse, bei denen der Großteil der Energieübertragung stattfindet.[18] “Diese Auslassungen und Probleme geben Anlass zur Sorge, aber nach heutigem Kenntnisstand ergeben sich keine unüberwindlichen Schwierigkeiten.”[4]::669
Es gibt verschiedene Arten oder Kategorien von Nahrungsnetzen:
- Quellweb – ein oder mehrere Knoten, alle ihre Raubtiere, alle Lebensmittel, die diese Raubtiere essen, und so weiter.
- Waschbecken – ein oder mehrere Knoten, alle ihre Beute, alle Lebensmittel, die diese Beute essen, und so weiter.
- Community (oder Verbundenheit) Web – eine Gruppe von Knoten und alle Verbindungen, wer wen isst.
- Energiefluss Web – quantifizierte Energieflüsse zwischen Knoten entlang der Verbindungen zwischen einer Ressource und einem Verbraucher.[4][40]
- Paläoökologisches Netz – ein Netz, das Ökosysteme aus dem Fossilienbestand rekonstruiert.[58]
- Funktionales Web – betont die funktionale Bedeutung bestimmter Verbindungen mit starker Interaktionsstärke und größerem Einfluss auf die Organisation der Gemeinschaft, mehr als Energieflusswege. Funktionale Netze haben Kompartimente, die Untergruppen im größeren Netzwerk sind, in denen es unterschiedliche Dichten und Stärken der Wechselwirkung gibt.[38][59] Funktionale Netze betonen, dass “die Bedeutung jeder Bevölkerung für die Aufrechterhaltung der Integrität einer Gemeinschaft sich in ihrem Einfluss auf die Wachstumsraten anderer Bevölkerungsgruppen widerspiegelt”.[40]::511
Innerhalb dieser Kategorien können Nahrungsnetze nach den verschiedenen Arten der untersuchten Ökosysteme weiter organisiert werden. Zum Beispiel menschliche Nahrungsnetze, landwirtschaftliche Nahrungsnetze, schädliche Nahrungsnetze, marine Nahrungsnetze, aquatische Nahrungsnetze, Bodennahrungsnetze, arktische (oder polare) Nahrungsnetze, terrestrische Nahrungsnetze und mikrobielle Nahrungsnetze. Diese Charakterisierungen stammen aus dem Ökosystemkonzept, bei dem davon ausgegangen wird, dass die untersuchten Phänomene (Wechselwirkungen und Rückkopplungsschleifen) ausreichen, um Muster innerhalb von Grenzen wie dem Rand eines Waldes, einer Insel, einer Küste oder einer anderen ausgeprägten physikalischen Eigenschaft zu erklären.[60][61][62]
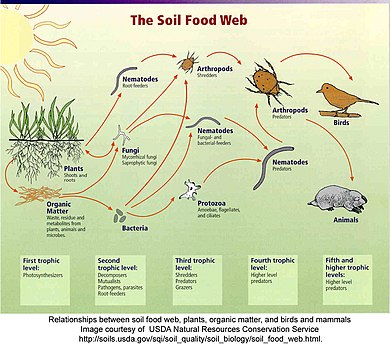
Detrital Web[edit]
In einem Detritalnetz werden pflanzliche und tierische Stoffe von Zersetzern, z. B. Bakterien und Pilzen, abgebaut und wandern zu Detritivoren und dann zu Fleischfressern.[63] Es gibt oft Beziehungen zwischen dem Detritalnetz und dem Weidegewebe. Pilze, die von Zersetzern im Schuttnetz produziert werden, werden zu einer Nahrungsquelle für Hirsche, Eichhörnchen und Mäuse im Weidenetz. Regenwürmer, die von Rotkehlchen gefressen werden, sind Detritivoren, die verfallende Blätter verzehren.[64]
“Detritus kann allgemein als jede Form von nicht lebender organischer Substanz definiert werden, einschließlich verschiedener Arten von Pflanzengewebe (z. B. Laub, totes Holz, aquatische Makrophyten, Algen), tierischem Gewebe (Aas), toten Mikroben, Kot (Gülle, Mist) , Kotpellets, Guano, Frass) sowie Produkte, die von Organismen abgesondert, ausgeschieden oder ausgeschieden werden (z. B. extrazelluläre Polymere, Nektar, Wurzelexsudate und Sickerwasser, gelöste organische Stoffe, extrazelluläre Matrix, Schleim). Die relative Bedeutung von Diese Formen von Detritus variieren in Bezug auf Herkunft, Größe und chemische Zusammensetzung zwischen den Ökosystemen. “[48]::585
Quantitative Nahrungsnetze[edit]
Ökologen sammeln Daten zu trophischen Niveaus und Nahrungsnetzen, um Parameter statistisch zu modellieren und mathematisch zu berechnen, wie sie beispielsweise bei anderen Arten der Netzwerkanalyse (z. B. Graphentheorie) verwendet werden, um emergente Muster und Eigenschaften zu untersuchen, die zwischen Ökosystemen geteilt werden. Es gibt verschiedene ökologische Dimensionen, die kartiert werden können, um kompliziertere Nahrungsnetze zu schaffen, darunter: Artenzusammensetzung (Art der Arten), Reichtum (Anzahl der Arten), Biomasse (Trockengewicht von Pflanzen und Tieren), Produktivität (Umwandlungsraten von Energie und Nährstoffe in Wachstum) und Stabilität (Nahrungsnetze im Laufe der Zeit). Ein Nahrungsnetzdiagramm, das die Artenzusammensetzung veranschaulicht, zeigt, wie Veränderungen bei einer einzelnen Art viele andere direkt und indirekt beeinflussen können. Mikrokosmosstudien werden verwendet, um die Erforschung des Nahrungsnetzes in halbisolierten Einheiten wie kleinen Quellen, verfallenden Baumstämmen und Laborexperimenten mit Organismen zu vereinfachen, die sich schnell vermehren, wie Daphnien, die sich von Algen ernähren, die unter kontrollierten Bedingungen in Wasserkrügen gezüchtet werden.[37][65]
Während die Komplexität realer Nahrungsnetzverbindungen schwer zu entschlüsseln ist, haben Ökologen mathematische Modelle in Netzwerken als ein unschätzbares Werkzeug angesehen, um Einblicke in die Struktur, Stabilität und Gesetze des Verhaltens von Nahrungsnetzen im Verhältnis zu beobachtbaren Ergebnissen zu erhalten. “Die Theorie des Nahrungsnetzes dreht sich um die Idee der Konnektivität.”[66]::1648 Quantitative Formeln vereinfachen die Komplexität der Nahrungsnetzstruktur. Die Anzahl der trophischen Links (tL.) wird beispielsweise in einen Verbindungswert umgewandelt:
- ,

wobei S (S-1) / 2 die maximale Anzahl von binären Verbindungen zwischen S-Spezies ist.[66] “Konnektivität (C) ist der Bruchteil aller möglichen Verbindungen, die realisiert werden (L / S.2) und stellt ein Standardmaß für die Komplexität des Nahrungsnetzes dar … “[67]::12913 Der Abstand (d) zwischen jedem Artenpaar in einem Netz wird gemittelt, um den mittleren Abstand zwischen allen Knoten in einem Netz (D) zu berechnen.[67] und multipliziert mit der Gesamtzahl der Verbindungen (L), um die Verbindungsdichte (LD) zu erhalten, die durch skalenabhängige Variablen wie den Artenreichtum beeinflusst wird. Diese Formeln sind die Grundlage für den Vergleich und die Untersuchung der Natur nicht zufälliger Muster in der Struktur von Nahrungsnetzwerken zwischen vielen verschiedenen Arten von Ökosystemen.[67][68]
Skalierungsgesetze, Komplexität, Chaos und Musterkorrelate sind gemeinsame Merkmale, die der Struktur des Nahrungsnetzes zugeschrieben werden.[69][70]
Komplexität und Stabilität[edit]
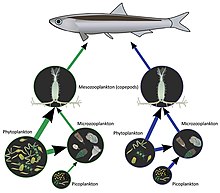
Nahrungsnetze sind äußerst komplex. Komplexität ist ein Maß für eine zunehmende Anzahl von Permutationen und ein metaphorischer Begriff, der die geistige Unlösbarkeit oder die Grenzen unbegrenzter algorithmischer Möglichkeiten vermittelt. In der Terminologie des Nahrungsnetzes ist Komplexität ein Produkt der Anzahl der Arten und der Konnektivität.[71][72][73] Konnektivität ist “der Bruchteil aller möglichen Verbindungen, die in einem Netzwerk realisiert werden”.[74]::12917 Diese Konzepte wurden abgeleitet und angeregt durch den Vorschlag, dass Komplexität zu Stabilität in Nahrungsnetzen führt, beispielsweise durch die Erhöhung der Anzahl trophischer Niveaus in artenreicheren Ökosystemen. Diese Hypothese wurde durch mathematische Modelle in Frage gestellt, die etwas anderes vorschlagen, aber nachfolgende Studien haben gezeigt, dass die Prämisse in realen Systemen gilt.[71][68]
Auf verschiedenen Ebenen in der Hierarchie des Lebens, wie beispielsweise der Stabilität eines Nahrungsnetzes, “bleibt die gleiche Gesamtstruktur trotz eines ständigen Flusses und Wechsels der Komponenten erhalten.”[75]::476 Je weiter ein lebendes System (z. B. ein Ökosystem) vom Gleichgewicht abweicht, desto komplexer ist es.[75] Komplexität hat in den Biowissenschaften und im öffentlichen Raum mehrere Bedeutungen, die ihre Anwendung als präzisen Begriff für analytische Zwecke in der Wissenschaft verwechseln.[73][76] Die Komplexität in den Biowissenschaften (oder die Biokomplexität) wird definiert durch die “Eigenschaften, die sich aus dem Zusammenspiel von Verhaltens-, biologischen, physischen und sozialen Interaktionen ergeben, die lebende Organismen, einschließlich Menschen, beeinflussen, aufrechterhalten oder von ihnen verändert werden”.[77]::1018
Aus der Untersuchung der Komplexität von Nahrungsnetzen sind mehrere Konzepte hervorgegangen. Die Komplexität erklärt viele Prinzipien, die sich auf Selbstorganisation, Nichtlinearität, Interaktion, kybernetisches Feedback, Diskontinuität, Entstehung und Stabilität in Nahrungsnetzen beziehen. Verschachtelung wird zum Beispiel definiert als “ein Interaktionsmuster, in dem Spezialisten mit Arten interagieren, die perfekte Teilmengen der Arten bilden, mit denen Generalisten interagieren”.[78]::575 “- das heißt, die Ernährung der am meisten spezialisierten Arten ist eine Teilmenge der Ernährung der nächst allgemeineren Arten, und ihre Ernährung ist eine Teilmenge der nächst allgemeineren Arten, und so weiter.”[79] Bis vor kurzem wurde angenommen, dass Nahrungsnetze wenig verschachtelte Strukturen haben, aber empirische Beweise zeigen, dass viele veröffentlichte Netze in ihrer Zusammenstellung verschachtelte Unternetze haben.[80]
Nahrungsnetze sind komplexe Netzwerke. Als Netzwerke weisen sie ähnliche strukturelle Eigenschaften und mathematische Gesetze auf, die zur Beschreibung anderer komplexer Systeme verwendet wurden, wie z. B. kleine Welt und skalierungsfreie Eigenschaften. Das Attribut “Kleine Welt” bezieht sich auf die vielen lose verbundenen Knoten, die nicht zufällige dichte Häufung einiger Knoten (dh trophische oder Keystone-Arten in der Ökologie) und die geringe Pfadlänge im Vergleich zu einem regulären Gitter.[74][81] “Ökologische Netzwerke, insbesondere wechselseitige Netzwerke, sind im Allgemeinen sehr heterogen und bestehen aus Gebieten mit geringen Verbindungen zwischen Arten und unterschiedlichen Gebieten eng verbundener Arten. Diese Regionen mit hoher Verbindungsdichte werden häufig als Cliquen, Hubs, Kompartimente, zusammenhängende Untergruppen bezeichnet , oder Module … Innerhalb von Nahrungsnetzen, insbesondere in aquatischen Systemen, scheint die Verschachtelung mit der Körpergröße in Zusammenhang zu stehen, da die Ernährung kleinerer Raubtiere tendenziell verschachtelte Teilmengen der Ernährung größerer Raubtiere ist (Woodward & Warren 2007; YvonDurocher et al. 2008) ) und phylogenetische Einschränkungen, bei denen verwandte Taxa aufgrund ihrer gemeinsamen Evolutionsgeschichte verschachtelt sind, sind ebenfalls offensichtlich (Cattin et al. 2004). “[82]::257 “Kompartimente in Nahrungsnetzen sind Untergruppen von Taxa, in denen viele starke Wechselwirkungen innerhalb der Untergruppen und wenige schwache Wechselwirkungen zwischen den Untergruppen auftreten. Theoretisch erhöhen Kompartimente die Stabilität in Netzwerken wie Nahrungsnetzen.”[59]
Nahrungsnetze sind auch insofern komplex, als sie sich saisonal und geografisch in ihrer Größe ändern. Die Bestandteile von Nahrungsnetzen, einschließlich Organismen und Mineralstoffen, überschreiten die Grenzen der Ökosystemgrenzen. Dies hat zu dem Konzept oder Studienbereich geführt, der als grenzüberschreitende Subvention bekannt ist.[60][61] “Dies führt zu Anomalien wie Berechnungen des Nahrungsnetzes, bei denen festgestellt wird, dass ein Ökosystem die Hälfte eines Top-Fleischfressers unterstützen kann, ohne anzugeben, an welchem Ende.”[62] Beim Vergleich verschiedener Arten von ökologischen Nahrungsnetzen, wie terrestrischen und aquatischen Nahrungsnetzen, wurden jedoch echte Unterschiede in Struktur und Funktion festgestellt.[83]
Geschichte der Nahrungsnetze[edit]
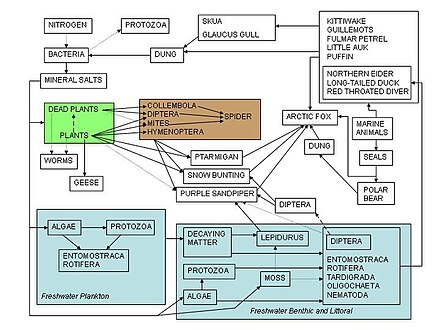
Nahrungsnetze dienen als Rahmen, um Ökologen dabei zu helfen, das komplexe Netzwerk von Wechselwirkungen zwischen Arten zu organisieren, die in der Natur und auf der ganzen Welt beobachtet werden. Eine der frühesten Beschreibungen einer Nahrungskette wurde von einem mittelalterlichen afro-arabischen Gelehrten namens Al-Jahiz beschrieben: “Kurz gesagt, alle Tiere können nicht ohne Nahrung existieren, und das Jagdtier kann auch nicht entkommen, wenn es seinerseits gejagt wird.”[84]::143 Die früheste grafische Darstellung eines Nahrungsnetzes stammt von Lorenzo Camerano aus dem Jahr 1880, gefolgt von denen von Pierce und Kollegen aus dem Jahr 1912 und Victor Shelford aus dem Jahr 1913.[85][86] Zwei Nahrungsnetze über Hering wurden von Victor Summerhayes und Charles Elton hergestellt[87] und Alister Hardy[88] 1923 und 1924. Charles Elton war in seinem klassischen Buch “Animal Ecology” von 1927 Pionier des Konzepts der Nahrungszyklen, Nahrungsketten und Nahrungsgrößen. Eltons “Nahrungszyklus” wurde in einem nachfolgenden ökologischen Text durch “Nahrungsnetz” ersetzt.[89] Nach Charles Eltons Verwendung von Nahrungsnetzen in seiner Synthese von 1927[90] Sie wurden zu einem zentralen Konzept im Bereich der Ökologie. Elton[89] organisierte Arten in funktionelle Gruppen, die 1942 die Grundlage für das trophische Klassifikationssystem in Raymond Lindemans klassischem und wegweisendem Papier über trophische Dynamik bildeten.[17][38][91] Der Begriff eines Nahrungsnetzes hat einen historischen Halt in den Schriften von Charles Darwin und seiner Terminologie, einschließlich einer “verschränkten Bank”, eines “Netzes des Lebens”, eines “Netzes komplexer Beziehungen” und in Bezug auf die Zersetzungsaktionen von Regenwürmern, die er hat sprach über “die fortgesetzte Bewegung der Teilchen der Erde”. Noch früher, 1768, beschrieb John Bruckner die Natur als “ein fortbestehendes Netz des Lebens”.[4][92][93][94]
Das Interesse an Nahrungsnetzen nahm nach Robert Paines experimenteller und deskriptiver Untersuchung der Gezeitenküste zu[95] Dies deutet darauf hin, dass die Komplexität des Nahrungsnetzes der Schlüssel zur Erhaltung der Artenvielfalt und der ökologischen Stabilität war. Viele theoretische Ökologen, darunter Sir Robert May[96] und Stuart Pimm,[97] wurden durch diese Entdeckung und andere veranlasst, die mathematischen Eigenschaften von Nahrungsnetzen zu untersuchen.
Siehe auch[edit]
Verweise[edit]
- ^ Nowak, ME; Beulig, F.; von Fischer, J.; Muhr, J.; Küsel, K.; Trumbore, SE (2015). “Die autotrophe Fixierung von geogenem CO2 durch Mikroorganismen trägt zur Bildung organischer Stoffe im Boden bei und verändert die Isotopensignaturen in einer Feuchtgebietsmofette.” (PDF). Biogeowissenschaften. Copernicus Publications (veröffentlicht am 08.12.2015). 12 (23): 7169–7183. doi:10.5194 / bg-12-7169-2015. Abgerufen 2019-10-01.
- ^ Kormondy, EJ (1996). Konzepte der Ökologie (4. Aufl.). New Jersey: Prentice-Hall. p. 559. ISBN 978-0-13-478116-7.
- ^ ein b Proulx, SR; Promislow, DEL; Phillips, PC (2005). “Netzwerkdenken in Ökologie und Evolution” (PDF). Trends in Ökologie und Evolution. 20 (6): 345–353. doi:10.1016 / j.tree.2005.04.004. PMID 16701391. Archiviert von das Original (PDF) am 15.08.2011.
- ^ ein b c d e f G Pimm, SL; Lawton, JH; Cohen, JE (1991). “Nahrungsnetzmuster und ihre Folgen” (PDF). Natur. 350 (6320): 669–674. Bibcode:1991Natur.350..669P. doi:10.1038 / 350669a0. S2CID 4267587. Archiviert von das Original (PDF) am 2010-06-10.
- ^ ein b c d e f Odum, EP; Barrett, GW (2005). Grundlagen der Ökologie (5. Aufl.). Brooks / Cole, ein Teil von Cengage Learning. ISBN 978-0-534-42066-6. Archiviert von das Original am 20.08.2011.
- ^ ein b Benke, AC (2010). “Sekundärproduktion”. Naturerziehungswissen. 1 (8): 5.
- ^ Allesina, S.; Alonso, D.; Pascual, M. (2008). “Ein allgemeines Modell für die Struktur des Nahrungsnetzes” (PDF). Wissenschaft. 320 (5876): 658–661. Bibcode:2008Sci … 320..658A. doi:10.1126 / science.1156269. PMID 18451301. S2CID 11536563. Archiviert von das Original (PDF) am 28.09.2011.
- ^ Azam, F.; Fenche, T.; Field, JG; Gra, JS; Meyer-Reil, LA; Thingstad, F. (1983). “Die ökologische Rolle von Wassersäulenmikroben im Meer” (PDF). Mar. Ecol. Prog. Ser. 10: 257–263. Bibcode:1983MEPS … 10..257A. doi:10.3354 / meps010257.
- ^ Uroz, S.; Calvarus, C.; Turpault, M.; Frey-Klett, P. (2009). “Mineralische Verwitterung durch Bakterien: Ökologie, Akteure und Mechanismen” (PDF). Trends in der Mikrobiologie. 17 (8): 378–387. doi:10.1016 / j.tim.2009.05.004. PMID 19660952.[permanent dead link]
- ^ Williams, RJ; Martinez, ND (2000). “Einfache Regeln ergeben komplexe Nahrungsnetze” (PDF). Natur. 404 (6774): 180–183. doi:10.1038 / 35004572. PMID 10724169. S2CID 205004984.
- ^ Post, DM (2002). “Die lange und kurze Länge der Nahrungskette” (PDF). Trends in Ökologie und Evolution. 17 (6): 269–277. doi:10.1016 / S0169-5347 (02) 02455-2. Archiviert von das Original (PDF) am 28.07.2011.
- ^ Tavares-Cromar, AF; Williams, DD (1996). “Die Bedeutung der zeitlichen Auflösung bei der Analyse von Nahrungsnetzen: Beweise aus einem Detritus-basierten Strom” (PDF). Ökologische Monographien. 66 (1): 91–113. doi:10.2307 / 2963482. hdl:1807/768. JSTOR 2963482.
- ^ ein b Pimm, SL (1979). “Die Struktur von Nahrungsnetzen” (PDF). Theoretische Populationsbiologie. 16 (2): 144–158. doi:10.1016 / 0040-5809 (79) 90010-8. PMID 538731. Archiviert von das Original (PDF) am 27.09.2011.
- ^ ein b Cousins, S. (1985-07-04). “Ökologen bauen wieder Pyramiden”. Neuer Wissenschaftler. 1463: 50–54.
- ^ McCann, K. (2007). “Schutz der Biostruktur” (PDF). Natur. 446 (7131): 29. Bibcode:2007Natur.446 … 29M. doi:10.1038 / 446029a. PMID 17330028. S2CID 4428058. Archiviert von das Original (PDF) am 22.07.2011.
- ^ ein b Thompson, RM; Hemberg, M.; Starzomski, BM; Shurin, JB (März 2007). “Trophäen und trophische Verwicklungen: Die Verbreitung von Allesfressern in echten Nahrungsnetzen” (PDF). Ökologie. 88 (3): 612–617. doi:10.1890 / 05-1454. PMID 17503589. Archiviert von das Original (PDF) am 15.08.2011.
- ^ ein b c Lindeman, RL (1942). “Der trophisch-dynamische Aspekt der Ökologie” (PDF). Ökologie. 23 (4): 399–417. doi:10.2307 / 1930126. JSTOR 1930126.
- ^ ein b Hairston, NG; Hairston, NG (1993). “Ursache-Wirkungs-Beziehungen im Energiefluss, in der trophischen Struktur und in interspezifischen Wechselwirkungen” (PDF). Der amerikanische Naturforscher. 142 (3): 379–411. doi:10.1086 / 285546. Archiviert von das Original (PDF) am 20.07.2011.
- ^ Fretwell, SD (1987). “Dynamik der Nahrungskette: Die zentrale Theorie der Ökologie?” (PDF). Oikos. 50 (3): 291–301. doi:10.2307 / 3565489. JSTOR 3565489. Archiviert von das Original (PDF) am 28.07.2011.
- ^ Polis, GA; Strong, DR (1996). “Komplexität des Nahrungsnetzes und Dynamik der Gemeinschaft” (PDF). Der amerikanische Naturforscher. 147 (5): 813–846. doi:10.1086 / 285880. S2CID 85155900.
- ^ Hoekman, D. (2010). “Kopf hochdrehen: Die Temperatur beeinflusst die relative Bedeutung von Top-Down- und Bottom-Up-Effekten” (PDF). Ökologie. 91 (10): 2819–2825. doi:10.1890 / 10-0260.1. PMID 21058543.
- ^ Schmitz, ABl. (2008). “Pflanzenfresser vom Individuum zum Ökosystem”. Jahresrückblick auf Ökologie, Evolution und Systematik. 39: 133–152. doi:10.1146 / annurev.ecolsys.39.110707.173418. S2CID 86686057.
- ^ Tscharntke, T.; Hawkins, B., A., Hrsg. (2002). Interaktionen auf multitrophischer Ebene. Cambridge: Cambridge University Press. p. 282. ISBN 978-0-521-79110-6.
- ^ Polis, GA; et al. (2000). “Wann ist eine trophische Kaskade eine trophische Kaskade?” (PDF). Trends in Ökologie und Evolution. 15 (11): 473–5. doi:10.1016 / S0169-5347 (00) 01971-6. PMID 11050351.
- ^ Sterner, RW; Klein, GE; Hood, JM “Die Erhaltung der Masse”. Naturerziehungswissen. 2 (1): 11.
- ^ Odum, HT (1988). “Selbstorganisation, Transformation und Information”. Wissenschaft. 242 (4882): 1132–1139. Bibcode:1988Sci … 242.1132O. doi:10.1126 / science.242.4882.1132. JSTOR 1702630. PMID 17799729. S2CID 27517361.
- ^ Odum, EP (1968). “Energiefluss in Ökosystemen: Ein historischer Rückblick”. Amerikanischer Zoologe. 8 (1): 11–18. doi:10.1093 / icb / 8.1.11.
- ^ Mann, KH (1988). “Produktion und Verwendung von Detritus in verschiedenen Süßwasser-, Flussmündungs- und Küstenmeerökosystemen” (PDF). Limnol. Oceanogr. 33 (2): 910–930. doi:10.4319 / lo.1988.33.4_part_2.0910. Archiviert von das Original (PDF) am 25.04.2012.
- ^ ein b Koijman, SALM; Andersen, T.; Koo, BW (2004). “Dynamische Energiebudgetdarstellungen stöchiometrischer Einschränkungen der Bevölkerungsdynamik” (PDF). Ökologie. 85 (5): 1230–1243. doi:10.1890 / 02-0250.
- ^ Anderson, KH; Beyer, JE; Lundberg, P. (2009). “Trophäe und individuelle Effizienz größenstrukturierter Gemeinschaften”. Proc Biol Sci. 276 (1654): 109–114. doi:10.1098 / rspb.2008.0951. PMC 2614255. PMID 18782750.
- ^ Benke, AC (2011). “Sekundärproduktion, quantitative Nahrungsnetze und trophische Position”. Naturerziehungswissen. 2 (2): 2.
- ^ Spellman, Frank R. (2008). Die Wissenschaft vom Wasser: Konzepte und Anwendungen. CRC Drücken Sie. p. 165. ISBN 978-1-4200-5544-3.
- ^ Kent, Michael (2000). Fortgeschrittene Biologie. Oxford University Press US. p. 511. ISBN 978-0-19-914195-1.
- ^ Kent, Michael (2000). Fortgeschrittene Biologie. Oxford University Press US. p. 510. ISBN 978-0-19-914195-1.
- ^ ein b Post, DM (1993). “Die lange und kurze Länge der Nahrungskette”. Trends in Ökologie und Evolution. 17 (6): 269–277. doi:10.1016 / S0169-5347 (02) 02455-2.
- ^ Odum, EP; Barrett, GW (2005). Grundlagen der Ökologie. Brooks Cole. p. 598. ISBN 978-0-534-42066-6.[permanent dead link]
- ^ ein b Worm, B.; Duffy, JE (2003). “Biodiversität, Produktivität und Stabilität in echten Nahrungsnetzen”. Trends in Ökologie und Evolution. 18 (12): 628–632. doi:10.1016 / j.tree.2003.09.003.
- ^ ein b c Paine, RT (1980). “Nahrungsnetze: Verknüpfung, Interaktionsstärke und Gemeinschaftsinfrastruktur”. Zeitschrift für Tierökologie. 49 (3): 666–685. doi:10.2307 / 4220. JSTOR 4220. S2CID 55981512.
- ^ Raffaelli, D. (2002). “Von Elton zur Mathematik und wieder zurück”. Wissenschaft. 296 (5570): 1035–1037. doi:10.1126 / science.1072080. PMID 12004106. S2CID 177263265.
- ^ ein b c Rickleffs, Robert, E. (1996). Die Ökonomie der Natur. University of Chicago Press. p. 678. ISBN 978-0-7167-3847-3.
- ^ Whitman, WB; Coleman, DC; Wieb, WJ (1998). “Prokaryoten: Die unsichtbare Mehrheit”. Proc. Natl. Acad. Sci. Vereinigte Staaten von Amerika. 95 (12): 6578–83. Bibcode:1998PNAS … 95.6578W. doi:10.1073 / pnas.95.12.6578. PMC 33863. PMID 9618454.
- ^ Groombridge, B.; Jenkins, M. (2002). Weltatlas der biologischen Vielfalt: Die lebenden Ressourcen der Erde im 21. Jahrhundert. World Conservation Monitoring Center, Umweltprogramm der Vereinten Nationen. ISBN 978-0-520-23668-4.
- ^ Spellman, Frank R. (2008). Die Wissenschaft vom Wasser: Konzepte und Anwendungen. CRC Drücken Sie. p. 167. ISBN 978-1-4200-5544-3.
- ^ Wang, H.; Morrison, W.; Singh, A.; Weiss, H. (2009). “Modellierung invertierter Biomassepyramiden und Schutzhütten in Ökosystemen” (PDF). Ökologische Modellierung. 220 (11): 1376–1382. doi:10.1016 / j.ecolmodel.2009.03.005. Archiviert von das Original (PDF) am 07.10.2011.
- ^ Pomeroy, LR (1970). “Die Strategie des Mineralienkreislaufs”. Jahresrückblick auf Ökologie und Systematik. 1: 171–190. doi:10.1146 / annurev.es.01.110170.001131. JSTOR 2096770.
- ^ Elser, JJ; Fagan, WF; Donno, RF; Dobberfuhl, DR; Folarin, A.; Huberty, A.; et al. (2000). “Ernährungsbeschränkungen in terrestrischen und Süßwasser-Nahrungsnetzen” (PDF). Natur. 408 (6812): 578–580. doi:10.1038 / 35046058. PMID 11117743. S2CID 4408787.[permanent dead link]
- ^ Koch, PL; Fox-Dobbs, K.; Newsom, SD Diet, GP; Flessa, KW (Hrsg.). “Die Isotopenökologie fossiler Wirbeltiere und die Paläobiologie des Naturschutzes” (PDF). Die Papiere der Paläontologischen Gesellschaft. 15: 95–112.
- ^ ein b Moore, JC; Berlow, EL; Coleman, DC; de Ruiter, PC; Dong, Q.; Hastings, A.; et al. (2004). “Detritus, trophische Dynamik und Biodiversität”. Ökologie-Briefe. 7 (7): 584–600. doi:10.1111 / j.1461-0248.2004.00606.x. S2CID 2635427.
- ^ HA, Lowenstam (1981). “Von Organismen gebildete Mineralien”. Wissenschaft. 211 (4487): 1126–1131. Bibcode:1981Sci … 211.1126L. doi:10.1126 / science.7008198. JSTOR 1685216. PMID 7008198. S2CID 31036238.
- ^ Warren, LA; Kauffman, ME (2003). “Mikrobielle Geoingenieure”. Wissenschaft. 299 (5609): 1027–1029. doi:10.1126 / science.1072076. JSTOR 3833546. PMID 12586932. S2CID 19993145.
- ^ González-Muñoz, MT; Rodriguez-Navarro, C.; Martínez-Ruiz, F.; Arias, JM; Merroun, ML; Rodriguez-Gallego, M. (2010). “Bakterielle Biomineralisation: Neue Erkenntnisse aus Myxococcus-induzierten Mineralniederschlägen”. Geological Society, London, Sonderpublikationen. 336 (1): 31–50. Bibcode:2010GSLSP.336 … 31G. doi:10.1144 / SP336.3. S2CID 130343033.
- ^ Gonzalez-Acosta, B.; Bashan, Y.; Hernandez-Saavedra, NY; Ascencio, F.; De la Cruz-Agüero, G. (2006). “Saisonale Meerwassertemperatur als Hauptdeterminante für Populationen kultivierbarer Bakterien in den Sedimenten einer intakten Mangrove in einer ariden Region” (PDF). FEMS Mikrobiologie Ökologie. 55 (2): 311–321. doi:10.1111 / j.1574-6941.2005.00019.x. PMID 16420638.
- ^ DeAngelis, DL; Mulholland, PJ; Palumbo, AV; Steinman, AD; Huston, MA; Elwood, JW (1989). “Nährstoffdynamik und Stabilität des Nahrungsnetzes”. Jahresrückblick auf Ökologie und Systematik. 20: 71–95. doi:10.1146 / annurev.ecolsys.20.1.71. JSTOR 2097085.
- ^ Twiss, MR; Campbell, PGC; Auclair, J. (1996). “Regeneration, Recycling und trophischer Transfer von Spurenmetallen durch mikrobielle Nahrungsnetzorganismen in den pelagischen Oberflächengewässern des Eriesees” (PDF). Limnologie und Ozeanographie. 41 (7): 1425–1437. Bibcode:1996LimOc..41.1425T. doi:10.4319 / lo.1996.41.7.1425. Archiviert von das Original (PDF) am 25.04.2012.
- ^ May, RM (1988). “Wie viele Arten gibt es auf der Erde?” (PDF). Wissenschaft. 241 (4872): 1441–1449. Bibcode:1988Sci … 241.1441M. doi:10.1126 / science.241.4872.1441. PMID 17790039. S2CID 34992724. Archiviert von das Original (PDF) am 11.05.2013. Abgerufen 2011-06-13.
- ^ Beattie, A.; Ehrlich, P. (2010). “Das fehlende Glied beim Erhalt der biologischen Vielfalt”. Wissenschaft. 328 (5976): 307–308. Bibcode:2010Sci … 328..307B. doi:10.1126 / science.328.5976.307-c. PMID 20395493.
- ^ Ehrlich, PR; Pringle, RM (2008). “Kolloquiumspapier: Wohin geht die biologische Vielfalt von hier aus? Eine düstere Business-as-usual-Prognose und ein hoffnungsvolles Portfolio an Teillösungen.”. Verfahren der Nationalen Akademie der Wissenschaften. 105 (S1): 11579–11586. Bibcode:2008PNAS..10511579E. doi:10.1073 / pnas.0801911105. PMC 2556413. PMID 18695214.
- ^ ein b Dunne, JA; Williams, RJ; Martinez, ND; Wood, RA; Erwin, DH; Dobson, Andrew P. (2008). “Zusammenstellung und Netzwerkanalyse von kambrischen Nahrungsnetzen”. PLOS Biologie. 6 (4): e102. doi:10.1371 / journal.pbio.0060102. PMC 2689700. PMID 18447582.
- ^ ein b Krause, AE; Frank, KA; Mason, DM; Ulanowicz, RE; Taylor, WW (2003). “Fächer in Nahrungsnetzstruktur offenbart” (PDF). Natur. 426 (6964): 282–285. Bibcode:2003Natur.426..282K. doi:10.1038 / nature02115. hdl:2027.42 / 62960. PMID 14628050. S2CID 1752696.
- ^ ein b Bormann, FH; Likens, GE (1967). “Nährstoffkreislauf” (PDF). Wissenschaft. 155 (3761): 424–429. Bibcode:1967Sci … 155..424B. doi:10.1126 / science.155.3761.424. PMID 17737551. S2CID 35880562. Archiviert von das Original (PDF) am 27.09.2011.
- ^ ein b Polis, GA; Anderson, WB; Hold, RD (1997). “Auf dem Weg zu einer Integration von Landschafts- und Nahrungsnetzökologie: Die Dynamik räumlich subventionierter Nahrungsnetze” (PDF). Jahresrückblick auf Ökologie und Systematik. 28: 289–316. doi:10.1146 / annurev.ecolsys.28.1.289. hdl:1808/817. Archiviert von das Original (PDF) am 2011-10-02.
- ^ ein b O’Neil, RV (2001). “Ist es Zeit, das Ökosystemkonzept zu begraben? (Natürlich mit vollen militärischen Ehren!)” (PDF). Ökologie. 82 (12): 3275–3284. doi:10.1890 / 0012-9658 (2001) 082[3275:IITTBT]2.0.CO; 2. Archiviert von das Original (PDF) am 25.04.2012.
- ^ Gönenç, I. Ethem; Koutitonsky, Vladimir G.; Rashleigh, Brenda (2007). Bewertung des Schicksals und der Auswirkungen toxischer Stoffe auf die Wasserressourcen. Springer. p. 279. ISBN 978-1-4020-5527-0.
- ^ Gil Nonato C. Santos; Alfonso C. Danac; Jorge P. Ocampo (2003). E-Biologie II. Rex Buchhandlung. p. 58. ISBN 978-971-23-3563-1.
- ^ Elser, J.; Hayakawa, K.; Urabe, J. (2001). “Nährstoffbegrenzung verringert die Lebensmittelqualität für Zooplankton: Daphnienreaktion auf Seston-Phosphoranreicherung”. Ökologie. 82 (3): 898–903. doi:10.1890 / 0012-9658 (2001) 082[0898:NLRFQF]2.0.CO; 2.
- ^ ein b Paine, RT (1988). “Roadmaps von Interaktionen oder Kern für die theoretische Entwicklung?” (PDF). Ökologie. 69 (6): 1648–1654. doi:10.2307 / 1941141. JSTOR 1941141. Archiviert von das Original (PDF) am 28.07.2011.
- ^ ein b c Williams, RJ; Berlow, EL; Dunne, JA; Barabási, A.; Martinez, ND (2002). “Zwei Trennungsgrade in komplexen Nahrungsnetzen”. Verfahren der Nationalen Akademie der Wissenschaften. 99 (20): 12913–12916. Bibcode:2002PNAS … 9912913W. doi:10.1073 / pnas.192448799. PMC 130559. PMID 12235367.
- ^ ein b Banasek-Richter, C.; Bersier, LL; Cattin, M.; Baltensperger, R.; Gabriel, J.; Merz, Y.; et al. (2009). “Komplexität in quantitativen Nahrungsnetzen” (PDF). Ökologie. 90 (6): 1470–1477. doi:10.1890 / 08-2207.1. hdl:1969.1 / 178777. PMID 19569361. Archiviert von das Original (PDF) am 01.06.2011.
- ^ Riede, JO; Rall, BC; Banasek-Richter, C.; Navarrete, SA; Wieters, EA; Emmerson, MC; et al. (2010). Woodwoard, G. (Hrsg.). Skalierung von Nahrungsnetzeigenschaften mit Vielfalt und Komplexität über Ökosysteme hinweg (PDF). 42. Burlington: Akademische Presse. S. 139–170. ISBN 978-0-12-381363-3.
- ^ Briand, F.; Cohen, JE (1987). “Umweltkorrelate der Länge der Nahrungskette” (PDF). Wissenschaft. 238 (4829): 956–960. Bibcode:1987Sci … 238..956B. doi:10.1126 / science.3672136. PMID 3672136. Archiviert von das Original (PDF) am 25.04.2012.
- ^ ein b Neutel, A.; Heesterbeek, JAP; de Ruiter, PD (2002). “Stabilität in echten Nahrungsnetzen: Schwache Verbindung in langen Schleifen” (PDF). Wissenschaft. 295 (550): 1120–1123. Bibcode:2002Sci … 296.1120N. doi:10.1126 / science.1068326. hdl:1874/8123. PMID 12004131. S2CID 34331654. Archiviert von das Original (PDF) am 28.09.2011.
- ^ Leveque, C., ed. (2003). Ökologie: Vom Ökosystem zur Biosphäre. Wissenschaftsverlage. p. 490. ISBN 978-1-57808-294-0.
- ^ ein b Proctor, JD; Larson, BMH (2005). “Ökologie, Komplexität und Metapher” (PDF). BioScience. 55 (12): 1065–1068. doi:10.1641 / 0006-3568 (2005) 055[1065:ECAM]2.0.CO; 2. Archiviert von das Original (PDF) am 06.10.2011.
- ^ ein b Dunne, JA; Williams, RJ; Martinez, ND (2002). “Food-Web-Struktur und Netzwerktheorie: Die Rolle von Konnektivität und Größe”. Verfahren der Nationalen Akademie der Wissenschaften. 99 (20): 12917–12922. Bibcode:2002PNAS … 9912917D. doi:10.1073 / pnas.192407699. PMC 130560. PMID 12235364.
- ^ ein b Capra, F. (2007). “Komplexität und Leben”. Syst. Res. 24 (5): 475–479. doi:10.1002 / sres.848.
- ^ Peters, RH (1988). “Einige allgemeine Probleme für die Ökologie, die durch die Theorie des Nahrungsnetzes veranschaulicht werden”. Ökologie. 69 (6): 1673–1676. doi:10.2307 / 1941145. JSTOR 1941145.
- ^ Michener, WK; Baerwald, TJ; Firth, P.; Palmer, MA; Rosenberger, JL; Sandlin, EA; Zimmerman, H. (2001). “Biokomplexität definieren und enträtseln” (PDF). BioScience. 51 (12): 1018–1023. doi:10.1641 / 0006-3568 (2001) 051[1018:daub]2.0.co; 2. Archiviert von das Original (PDF) am 17.08.2011. Abgerufen 2011-07-04.
- ^ Bascompte, J.; Jordan, P. (2007). “Gegenseitige Netzwerke zwischen Pflanzen und Tieren: Die Architektur der biologischen Vielfalt” (PDF). Annu. Rev. Ecol. Evol. Syst. 38: 567–569. doi:10.1146 / annurev.ecolsys.38.091206.095818. Archiviert von das Original (PDF) am 25.10.2009.
- ^ Montoya, JM; Pimm, SL; Solé, RV (2006). “Ökologische Netzwerke und ihre Fragilität” (PDF). Natur. 442 (7100): 259–264. Bibcode:2006Natur.442..259M. doi:10.1038 / nature04927. PMID 16855581. S2CID 592403. Archiviert von das Original (PDF) am 06.07.2010.
- ^ Michio, K.; Kato, S.; Sakato, Y. (2010). “Nahrungsnetze werden mit verschachtelten Unterweben aufgebaut”. Ökologie. 91 (11): 3123–3130. doi:10.1890 / 09-2219.1. PMID 21141173.
- ^ Montoya, JM; Solé, RV (2002). “Kleine Weltmuster in Nahrungsnetzen” (PDF). Zeitschrift für Theoretische Biologie. 214 (3): 405–412. arXiv:cond-mat / 0011195. doi:10.1006 / jtbi.2001.2460. PMID 11846598. Archiviert von das Original (PDF) am 05.09.2011.
- ^ Montoya, JM; Blüthgen, N; Brown, L.; Dormann, CF; Edwards, F.; Figueroa, D.; et al. (2009). “Ökologische Netzwerke: jenseits von Nahrungsnetzen” (PDF). Zeitschrift für Tierökologie. 78 (1): 253–269. doi:10.1111 / j.1365-2656.2008.01460.x. PMID 19120606. Archiviert von das Original (PDF) am 16.09.2011.
- ^ Shurin, JB; Gruner, DS; Hillebrand, H. (2006). “Alles nass oder ausgetrocknet? Echte Unterschiede zwischen aquatischen und terrestrischen Nahrungsnetzen”. Proc. R. Soc. B.. 273 (1582): 1–9. doi:10.1098 / rspb.2005.3377. PMC 1560001. PMID 16519227.
- ^ Egerton, FN “Eine Geschichte der ökologischen Wissenschaften, Teil 6: Arabische Sprachwissenschaft: Ursprünge und zoologische Schriften” (PDF). Bulletin der Ecological Society of America. 83 (2): 142–146.
- ^ Egerton, FN (2007). “Nahrungsketten und Nahrungsnetze verstehen, 1700-1970”. Bulletin der Ecological Society of America. 88: 50–69. doi:10.1890 / 0012-9623 (2007) 88[50:UFCAFW]2.0.CO; 2.
- ^ Shelford, V. (1913). “Tiergemeinschaften im gemäßigten Amerika, wie in der Region Chicago dargestellt”. University of Chicago Press.
- ^ Summerhayes, VS; Elton, CS (1923). “Beiträge zur Ökologie Spitzbergens und der Bäreninsel”. Zeitschrift für Ökologie. 11 (2): 214–286. doi:10.2307 / 2255864. JSTOR 2255864.
- ^ Hardy, AC (1924). “Der Hering in Bezug auf seine belebte Umgebung. Teil 1. Die Ernährungs- und Ernährungsgewohnheiten des Herings unter besonderer Berücksichtigung der Ostküste Englands”. Fischereiuntersuchung London Serie II. 7 (3): 1–53.
- ^ ein b Elton, CS (1927). Tierökologie. London, UK.: Sidgwick und Jackson. ISBN 978-0-226-20639-4.
- ^ Elton CS (1927) Tierökologie. Neuauflage 2001. University of Chicago Press.
- ^ Allee, WC (1932). Tierleben und soziales Wachstum. Baltimore: Die Williams & Wilkins Company und Mitarbeiter.
- ^ Stauffer, RC (1960). “Ökologie in der langen Manuskriptversion von Darwins” Origin of Species “und Linnaeus ‘” Oeconomy of Nature ““”“. Proc. Am. Philos. Soc. 104 (2): 235–241. JSTOR 985662.
- ^ Darwin, CR (1881). “Die Bildung von Pflanzenschimmel durch die Einwirkung von Würmern mit Beobachtungen über ihre Gewohnheiten”. London: John Murray.
- ^ Worster, D. (1994). Naturökonomie: Eine Geschichte ökologischer Ideen (2. Aufl.). Cambridge University Press. p. 423. ISBN 978-0-521-46834-3.
- ^ Paine, RT (1966). “Komplexität des Nahrungsnetzes und Artenvielfalt”. Der amerikanische Naturforscher. 100 (910): 65–75. doi:10.1086 / 282400. S2CID 85265656.
- ^ Mai RM (1973) Stabilität und Komplexität in Modellökosystemen. Princeton University Press.
- ^ Pimm SL (1982) Food Webs, Chapman & Hall.
Weiterführende Literatur[edit]
Recent Comments