セグメンテーション遺伝子 – ウィキペディア
セグメンテーション遺伝子 昆虫の胚形成中のセグメントの数と内部組織を決定します。それらはモデル生物について研究されていました ショウジョウバエのメラノガスター 、特に、変異した飛行またはその胚の遺伝子を分析することにより、セグメンテーションまたは身体構造の奇形が示されます。遺伝子は、通常、その発見につながった突然変異に由来する名前を付けます。セグメンテーション遺伝子の遺伝子産物は、DNAおよび他の遺伝子に付着した調節タスクを備えたタンパク質であり、オンとオフのスイッチと同様に、これらは転写因子と呼ばれます。この目的のために、標的遺伝子には、タンパク質をコードする遺伝子断面の前にDNA鎖に配置された配列があり、それ自体が転写されていません。 DNA鎖のフロントエンドは「CIS」端(「トランス」と呼ばれる)と呼ばれるため、「CIS調節」セクションまたはCIS要素について話します。セグメンテーション遺伝子の転写因子は、調節カスケードに連続して切り替えられます。つまり、発達中の胚の位置に応じて、包括的で顕著なセグメンテーション遺伝子を早期に発音します。連続して配置されたストライプからのこのようなストリップパターンが発生し、特定のセグメンテーション遺伝子(またはそのようなグループ)がアクティブになります。このセルストライプは、体の後のセグメンテーションを好みます。体組織自体を構築する細胞には、セグメンテーション遺伝子を介して発達中の生物の位置に関する情報が与えられます。場所によっては、自分自身を成長させ、分割し、差別化するか、死ぬことができます(プログラムされた細胞死またはアポトーシス)。
後の分析では、動物界のセグメンテーション遺伝子の相同遺伝子が検査されたすべての生物で発生し、どこでも肉体の形成を組織することが示されています。これは、分割されていない生物の場合、非常に類似した類似の方法で行われます。セグメンテーション遺伝子のその他は、昆虫や節足動物でのみ顕著であり、いくつかは2つの流体(双pter)でさえ、個人はからのみです。 ショウジョウバエ 知られています。
洗礼の胚 ショウジョウバエ 卵細胞から外側の細胞層である胚盤胚葉によって発達し、中央の卵黄の供給を成長させます。芽版は爆風内で区別します最初の分裂ステップでは、細胞核のみが共有されますが、細胞質は細胞膜によって共有されていません。このような分割されていない包帯はSyncytiumと呼ばれます。形態学的には、この段階では認識される構造はありません。新興胚の前端と後端は同じように見えます。 1960年代と1970年代の実験は、身体の縦軸がこの段階ですでに決定されていることを示しています。卵がすでに母親によって与えられている縦軸を定義する組織化要因があることが判明しました。これは、前部または後部の卵極に濃縮された物質に基づいています。これらの遺伝子の変異は、深刻な奇形につながります。 B.体の前半分のシステムの代わりに、後半は2回です。対応する遺伝子産物(転写産物)は母親から来ているため、関連する遺伝子は 母体遺伝子 呼び出されました。したがって、氷の細胞質に移動した母体遺伝子の生成物は、胚の前端と後端を決定します。それらのタンパク質は、胚(勾配)に濃度勾配を形成します(勾配)。一部は最高濃度の上端で発生し、後端に発生します。その間のセルは、前面または背面に応じて、両方の異なる部分を含んでいます。母体遺伝子のタンパク質の濃度に応じて、他の遺伝子のグループが胚で活性化されます。 ギャップ (より頻繁に、英語、ギャップ遺伝子から採取)。命名は、これらの遺伝子の1つの突然変異に全身切片がないという事実に基づいています。で ショウジョウバエ 5つのギャップ遺伝子があります。各ギャップ遺伝子は、場所に応じて別の遺伝子を活性化するようになりました パアレゲルジェーン (Engl。PairRule Gene)。名前は、胚変異がセグメント数の半分しかない場合に基づいています。これにより、胚が7つのストリップで整理されます。カップル制御遺伝子は次のクラスを活性化します セグメントポラリゼーション – 遺伝子 。各ストリップは2つの基準に分かれています。その結果、製造されたフライの14のボディセグメントは教育を受けています。 (実際、この問題は少し複雑です。そのため、コールされたパラセグは教育を受けており、それぞれが隣のセグメントの端と後端の1つで構成されています)。
しかし、完成した生物では、これらのセグメントは形態学的に等しくありませんが、体切片(タグマタ)に分化します:頭、胴体(胸部)、腹部(腹部)。場所に応じて、異なるアタッチメント、およびアンテナ、脚などなどの他の臓器が訓練されているかどうかです。このアイデンティティは、別の遺伝子クラスによってセグメントに割り当てられます。 Hox-gene 。洗礼には8つのHox遺伝子があります。 HOX遺伝子の発現は、セグメントセグメントに正確にリンクされていません。一部の体セグメントは同じHox遺伝子を表現し、他の2つの遺伝子では2つが発現しています。
このモデル形成に関与する合計40〜50個の遺伝子が特定されており、それらのいくつかの役割はまだ不明です。母体および胚遺伝子も関与する同様のシグナル配列が、胚の背腹軸を決定します。 H.間違いなく上(裏側)および下(腹側)。これらの遺伝子は後に発見され、全体的にあまり知られていませんでした。
名前が示すように、母体の遺伝子は母体生物で活性です。あなたの遺伝子産物、主にRNA(転写産物)、めったにすでに完成していないタンパク質は、発達中の卵が与えられます。遺伝子尾側(CAD)とHunchback(HB)は二重の役割を持ち、胚自体の母体と後の両方で転写されます。体の縦軸の方向を指定する母体遺伝子の中で最も重要なのは、双子(BCD)です。バイコイドタンパク質は胚のフロントエンドを定義し、濃度に応じて、異なる幅のゾーンの他の遺伝子を逆方向に進行します。 CADとHBは、最初はほぼ均等に卵に分布しています。しかし、BCDはあなたの発現を阻害するため、それらは背中に蓄積します。 RNAとタンパク質の別のグループは、リアEipolで濃縮されています。
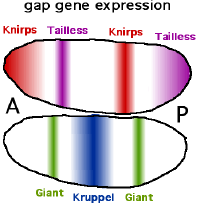

名前のギャップ(英語のギャップ遺伝子)は、これらの遺伝子の機能的故障がセグメンテーションのギャップ、身体セグメントの欠如につながるためです。 [初め] [2] 彼らは、フロント、ミディアム、リアエリアへの分割を担当しています。 [2] 現在11の知られているギャップには、巨人(GTまたはGAT)、ハンチバック(HB)、KNIRPS(KNI)、Krüppel(主に「Kruppel」)(KR)、Tailless(TLL)が含まれます。 CADとHBには二重の役割があります。それらは、母体(母体起源から)と胚自体の両方で表現されます。ギャップ遺伝子は当初、比較的広く発現し、後に自己組織化により右のストライプに限定されます。母体遺伝子の生成物と同様に、発現した転写因子は短時間のみ活性があります。パターンが開発され、次の手順が開始されるとすぐにアクティビティが終了します。これは、約2時間の開発の後に当てはまります。後の生物では、彼らはもはや役割を果たしていませんが、時には身体軸の決定における役割に関係なく、他のサンプル教育プロセスに関与することがあります。
ギャップ遺伝子の発現は、母体遺伝子とギャップ遺伝子の相互作用の組み合わせによって調節されます。さらに、これは細胞で役割を果たしますが、間には胴体(GATE)では役割を果たします。胴体は、卵殻内の物質によって活性化される膜貫通受容体をコードします。母体のBCDタンパク質は、体の前半分のHB遺伝子に切り替えられ、2つの鋭く離婚した半分が生じる(オールオアナッシング反応)。同時に、BCDはCADの転写を抑制し、CADタンパク質が体の後半でのみ発生するようにします。同様に、他のギャップ遺伝子は、胚の異なる点で1つまたは2つのストライプを形成します。このようにして、KRは主にBCDによってアクティブ化(予防)することにより、中心線の近くの領域で形成されますが、HBによって抑制(予防)されます。 KNIは、前面の端と背面のストリップで同等のメカニズムによって表されます。
通常、異なるボディセクションは、1つまたは2つのギャップタンパク質の組み合わせによって特徴付けられます。ギャップタンパク質は、一般に、背後に傾斜している最大の増加濃度に示され、その領域は多かれ少なかれ広く重複しています。
ギャップ遺伝子によって指定された胚性ストリップは、それぞれペアルール遺伝子の異なる組み合わせによって指定されます。カップルのルールジェーンは、均等なスキップ(Eve)とFushi Tarazu(FTZ)は、それぞれ7つのストリップで交互に表現されています。 Runt(Run)やHairy(H)などの他の遺伝子は、同様のパターンを示しています。したがって、ギャップ遺伝子の非周期的なストリップパターンは、周期的なパターンによってオーバーレイされます。ペアルールタンパク質の濃度は、細胞に応じて急激に離婚しており、ギャップ遺伝子のように重複しなくなります。ペアルール遺伝子の変異は、各2番目のセグメントの損失を引き起こすため、カップルルール遺伝子はロッドカウントまたは追放桁セグメントの形成を制御します。 [3]
セグメント極性遺伝子は、(パラ)セグメントの最終シーケンスとその極性の両方を照らします。 H.あなたのフロントエンドとリアエンド、しっかり。 Engrailed(S)セグメントの極性は、14のパラゼストリップの前面近くの狭いゾーンで表されます。ヘッジホッグ(HH)も同様のパターンを示しています。一方、遺伝子ウィングレス(WN)は、パラゼセグメントストリップの後端近くのストライプでアクティブになります。このパターンは、胚の生殖ストリップを同様のストライプに分割しますが、これは同期ではなく生成されますが、次々と生成されます。前述のセグメンテーション遺伝子とは対照的に、翼のある昆虫(Imago)に巻き込まれた場合、セグメント極性遺伝子は長い間アクティブのままです。セグメント極性遺伝子の活性により、セグメンテーションが完了します。開発の次の段階では、さまざまなセグメントのアイデンティティがさらに指定されています。これは何よりもHOX遺伝子のタスクです。特定のHOX遺伝子を活性化する場合、ギャップ遺伝子(セグメンテーションカスケードの後期段階とは対照的に、定期的ではありません)が重要な役割を果たします。
モデル生物について ショウジョウバエ 発見された基本スキームは、調査したすべての節足動物の基本的な特徴の後の研究で見つかりました。すべてのタイプについて、セグメント形成は、セグメント極性遺伝子のセグメントで構成されている生殖帯を介して、レギュレーションカスケードを介して実行されます。ただし、その前の開発の段階は、他の節足動物では詳細に異なります。これはZです。 B.ほとんどの節足動物は、ショウジョウバエとは異なり、シンシチウムで開発を開始しないこと。さらに、セグメントは多くの種に対して同時に作成されるのではなく、後端のセグメント層ゾーンの個体発生の過程で徐々に形成されます。これまでに検査された比較的少数の種では、これまでのところ、他の2つの流体(双pter)でしか見られていません(またはホモログ)。他の種では、母体の遺伝子も構造に関与していますが、その他は詳細に関与しています。遺伝子CADとNANO(NOS)は広まっているようです。ほとんどのセグメント形成遺伝子に対するオルソロジストまたは相同体遺伝子 ショウジョウバエ ほとんどの節足動物で発見されていますが、詳細な役割は異なるようです。さらに、他の調節パターンは、脊椎動物の体節のサンプル形成によりよく似たアークニドと千フィートにも見られました。
- Michael Akam(1987):ショウジョウバエ胚におけるメタメラパターンの分子基盤。開発101:1-22。
- Dmitri Papatsenko、Michael Levine(2011):ショウジョウバエギャップ遺伝子ネットワークは、2つの並列トグルスイッチで構成されています。 PLOS ONE 6(7):E21145。 doi: 10.1371/journal.pone.0021145
- Andrew D. Peel、Ariel D. Chipman、Michael Akam(2005):節足動物セグメンテーション:ショウジョウバエのパラダイムを超えて。 Nature Review Genetics doi: 10.1038/nrg1724
- ↑ C.R. Bartram et al。: ヒト – ネート診断:科学的基盤と社会的結果。 Springer、2000、ISBN 978-3540679455、S。30。
- ↑ a b ヤン・ヴォジェ、デビッド・ストーチ、スタニスラフ・ミフルカ: 進化:読書教科書。 Spectrum Akademischer Verlag、2009、ISBN 978-3827419750、p。230。
- ↑ Monica Hirsch-Kauffmann、Manfred Schweiger: 医師と科学者向けの生物学および分子医学。 そのeirmebuerg、2009、2009、Issbred 978-31376 pbox7aceship7、電話が返されました。254。
Recent Comments