Parasaurolophus – ウィキペディア
パラサウロロフス ハドロサウルス人のグループの恐竜の属であり、これは鳥類(鳥の盆地)の1つです。これらの動物の発見は、アルバータ州(カナダ)、ユタ州、ニューメキシコ州(米国)のアッパーチョーク(後期カンパニウム)の層から来ています。すべてのハドロサウルス人のように、草食動物はおそらく2本の足と4本の足を動かすことができました [2] そして、特徴的な歯のない「アヒルのくちばし」と顎に食事の歯がありました。最も印象的な機能は、ヘッドドレス、長い背中の骨鉛筆です。 パラサウロロフス 恐竜の最も知られている代表者の一人です。その顕著な外観のためです。

他のいくつかの人気のある科学とは対照的に、ハドロサウルスは比較的少なかった。 パラサウロロフス – デプスが見つかりました。つまり、スケルトンは完全にはわかっていません。のholotypusコピー Parasaurolophus Walker 長さは9.5 mと推定され、頭蓋骨と骨鉛筆とともに約1.6 mです。ホロタイプスのコピーの頭蓋骨と骨鉛筆 Parasaurolophus Tubicen 2 mを測定します。 [3] 生きている動物の重量は約2.5トンでした。 [4] 唯一の既知の前脚は、他のハドロサウルス菌と比較して比較的短いです。これにより、肩甲骨は短いですが広いです。太ももの骨(大腿骨) Parasaurolophus Walker 長さは103 cmで、その長さは非常に堅牢です。肩の領域と骨盤の骨も非常に困難でした。 [5] [6]
他のランボオサウリンのように パラサウロロフス 高いとげプロセス [5] これにより、背中と尾のアプローチが大幅に増加しました。とげのプロセスは、腰よりも最高点に達しました。から Parasaurolophus Walker 均一な結節のようなスケールを示すスキンプリントにも知られていますが、大きな構造はありません。 [7]

属の最も印象的な特徴は、頭蓋骨の骨鉛筆でした。これは、頭の上で後方に突き出ており、鼻の脚と中間松(前軸)から形成された長いチューブです。骨円錐はほとんど中空でしたが、チューブは鼻孔から端まで導かれ、曲線を作り、頭蓋骨に戻りました。これらのチューブ Parasaurolophus Walker 比較的単純でしたが、構造はそうでした Parasaurolophus Tubicen より複雑。このタイプでは、チューブは迷路の形の分割、崩壊、行き止まりを示しました。 [8]
のホロタイプスケルトン付き Parasaurolophus Walker 骨の鉛筆が背中に触れた時点にある椎骨プロセスで発見されました。動物が走っている間にこのノッチに骨ピンを置いた後、仮定がありました。 [9] ただし、この特性はおそらく病理学的であり、この個人の特別な特徴です。 [5] ウィリアム・パークス、動物の最初の説明、骨蛇の端から脊椎まで走るべき想定されたリーグまたは筋肉は、頭を支えるためにノッチングの領域の椎骨まで走るべきです。 [7] しかし、これはありそうもないようです [4] – しかし、このアイデアの残りは、骨鉛筆と背中の間のいくつかの再構成で緊張した皮膚フラップです。いくつかのイラストレーターのアイデアによると、この仮想の皮膚フラップと増加した背中は、動脈検出またはブライダル広告のためにカラフルに色付けされていました。 [十]

ハドロサウルス科内はそうです パラサウロロフス ハドロサウリネンの近くの親relativeだったとしても、ランボオサウリン人によって数えられます サウロロフス 間違い。 [11] 次の親relative パラサウロロフス たぶんだった Charonosaurus 中国北東部のアムール地域から。 Charonosaurus 同じような頭蓋骨があります パラサウロロフス ただし、骨の鉛筆は不完全に伝えられているだけです。これらの属の密接な関係は、頭蓋骨と脚の骨の共通特性(Synapomorphia)に付随しています。たとえば、ペアの額の脚は、骨のタップをサポートするためのプラットフォームを形成し、それは上側窓の一部に拡張されました。とは対照的に Charonosaurus 示した パラサウロロフス しかし、たとえば、頂点(頭頂)の部分の矢状コム。 [12番目] [13] パラサウロロフス と Charonosaurus パラサウロロフィニとしてランボオサウリナイ内で要約されており、他のすべてのランボオサウリンを含む姉妹グループのランベオサウリニと比較されています。 [14] [2] [15]
これまでのところ、3種類の属が パラサウロロフス 説明された:
タイプのタイプから Parasaurolophus Walker (Parks、1922)単一のスケルトンは、完全な頭蓋骨を持つアルバータ州から知られています。 [2] このタイプは異なります Parasaurolophus Tubicen 骨タップのシンプルなチューブ構造によって。とは対照的に Parasaurolophus cyrtocristatus 彼女は長い、曲がった骨のピンを見せました。他のすべてのランボオサウリンとは対照的に Parasaurolophus cyrtocristatus 、示した Parasaurolophus Walker さらに、前腕よりも前の首とより長い上腕の骨(上腕骨)の首が深くなります。 [15]
Parasaurolophus Tubicen (Wiman、1931)ニューメキシコ州のカートランド層からは、少なくとも3人の個人の発見を通じて知られています。 [2] それは最大の種でした、骨円錐により複雑なチューブを示しました Parasaurolophus Walker そして、より長くてまっすぐな骨円錐を持っていました Parasaurolophus cyrtocristatus 。 [8]
Parasaurolophus cyrtocristatus (オストロム、1961年)は、ニューメキシコ(フルーツランド層)とユタ州のカイパロウィッツ層でも発見されました。 3つの可能な発見が知られています。それは最小の方法であり、短くて丸い骨鉛筆が特徴でした。これは、それが女性または若い動物であるという仮説につながりました Parasaurolophus Tubicen 行動することができます。 [16] 最後に、このアイデアはトーマス・ウィリアムソンによって表されました。 Parasaurolophus cyrtocristatus サイズの約72% Parasaurolophus Tubicen ほぼ同じサイズの比率があります。これは、成虫と若い動物の間の他のランボオサウリナイでも疑われており、明確な性別の二型を訓練し始めています。 [16] しかし、この見解は最近の論文で拒否されました。 [2] [15] あるいは、他の2つのタイプは、 Parasaurolophus cyrtocristatus 、この種のサイズ偏差が必要です。 [15]
アルバータ州の恐竜地方公園の若い動物による脳の頭蓋骨は、既存の種に割り当てることができませんでした。 [13] アルバータからのさらに3つの発見もできるかもしれません パラサウロロフス 帰属します。 [17]
生息地 [ 編集 | ソーステキストを編集します ]

Parasaurolophus Walker アルバータ州の恐竜公園の形成から、豊かでよく文書化された恐竜動物の一部であるCeratopsier(Horndinosaur)の一部 セントロサウルス 、 スティラコサウルス と チャズモサウルス 、より多くのハドロサウルスのようです Prosaurolophus 、 グリプサウルス 、 コリトサウルス と ランベオサウルス 、アンキロサウリアル(装甲恐竜)のような エドモントニア と Euoplocephalus Tyrannosauridsと同様に ゴルゴサウルス 構成されています。 [18] この動物相の珍しい部分でした。 [19] 景観はおそらく川によって渡られ、振り出され、気候は西部の海路マリンの影響を受けました。 [17] 気候は今日のアルバータよりも暖かく、冬には霜が降りませんでしたが、湿った季節で乾燥していました。植物相は、葉の屋根を形成した針葉樹、シダ、木シダ、覆われた針葉樹で構成されていました。 [20]
一方、ニューメキシコ州のカートランド層の種は、大きな竜脚類とともに住んでいました アラモサウルス 、ハドロサウルー クリトサウルス 、dem ceratopsier Pentaceratops 、アンキロサウリエ 結節性の形状 、小さな野足原 saurornitholestes 以前に決定されていなかったティラノサウルスと同様に。 [18] カートランド層の堆積物は、西部の船路が撤回された後に作成された洪水レベルを表す可能性があります。針葉樹が植物相を支配し、Chasmosurine Ceratopier(角のある恐竜のグループ)は、ハドロサウルスよりも頻繁に現れたようです。 [21]
ダイエット [ 編集 | ソーステキストを編集します ]
ハドロサウルス科のメンバーとして パラサウロロフス 高度な顎が噛む動きを粉砕することを可能にする、大きくて4本足の2本足の有能な草食動物。噛むストリップは、数百本の歯が配置されているため、同時に数百本の歯が配置されていたため、まとめられた歯のバッテリーが形成されました。摩耗した歯は常に交換されていました。植物材料は最初にくちばしで摘み取られ、次に頬のようなマウスパートに保たれました。彼らはおそらく地面から4メートルまで放牧するかもしれません。 [2] 古生物学者のボブ・バッカーは、ハドロサウルス科の他のサブファミリーであるハドロサウルナエよりも狭い鼻を持っていると言いました。これは、ランボオサウリナエが広範囲にわたる、主にコンブレスのカウンターパートよりも選択的なフィーダーであるという理論につながりました。 [22]
骨タップ [ 編集 | ソーステキストを編集します ]
骨タップの機能を説明しようとする多数の仮説が設定されましたが、ほとんどは拒否されました。 [23] [4] 今日、いくつかの機能があります:視覚機能(タイプと性別の識別)、コミュニケーションの呼び出しのための共鳴体としての効果、および温度調節。しかし、進化の何時に最も重要だったかは明らかではありません。 [24]
[ 編集 | ソーステキストを編集します ]
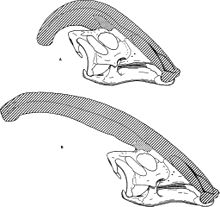
他のランボオサウリンと同様に、あなたもできます パラサウロロフス ヘッドの飾りは年齢とともに変化し、男性と女性(性別の二形性)で異なると仮定しました。
ジェームズ・ホプソンは、これらの違いに関連してランボオサウルスの装飾品を説明した最初の研究者の一人であり、それを疑った Parasaurolophus cyrtocristatus 、短い丸い骨円錐を持つ小さな種は女性でした。 [十] しかし、トーマス・ウィリアムソンは、それは若い動物だと言った Parasaurolophus Tubicen 。 [16] 他の研究者を参照してください Parasaurolophus cyrtocristatus 一方、独立した方法として。これらのアイデアはどれも大部分が受け入れられていません。合計6つの頭蓋骨と若い動物の頭蓋骨しか知らないので、それ以上の発見は明らかになります。
ウィリアムソンは、若い動物には性的成熟の直前に成長が加速した小さな装飾品があると疑っています。 [16] 以前は若い動物の脳の頭蓋骨に関する新しい研究 ランベオサウルス 起きたが、今 パラサウロロフス 割り当てられ、少年動物の小さな櫛に関する情報を提供します。発見は、ペアの正面脚(前頭)によって形成された小さな膨らみを示しています。同様の構造は、成体動物の化石からも知られていますが、そこははるかに大きいだけです。ここでは、前頭骨は骨毛布の基部をサポートするプラットフォームを形成します。 [13]
関数に関する反論された仮説 [ 編集 | ソーステキストを編集します ]

過去には、骨タップの機能を説明しようとした多数の仮説がありました。 [23] [4] 過去には、多くの研究者が仮説に行きましたが、それは1960年代まで広まっていましたが、ハドロサウルス人は水陸両用であり、水生ライフスタイルへの適応でした。たとえば、Alfred Sherwood Romer(1933)は、骨タップのシュノーケルのような機能を提案しました [25] 、Martin Wilfarth(1947)はトランクのブラケットを考えていましたが、それは呼吸して食べ物を集めるのに役立つはずでした。 [26] チャールズ・M・スターンバーグ(1935)は、それが空気で肺から水を遠ざけるための施設であると仮定しました [27] 、およびNed Colbert(1945)が疑われる、中空の骨円錐は、より長いダイブを可能にする空気貯水池として機能します。 [28]
他の仮定はより物理的でした。前述のように、最初の記述子ウィリアムパークは、骨の端を後椎骨に接続する靭帯を信じていました – 頭を安定させました。 [7] オセニオ・アベルは骨の鉛筆を武器と見なしました。 パラサウロロフス 使われた [29] 、そしてアンジェラ・ミルナーは、今日のカスアーが彼のヘルメットで行っているように、茂みの中で植生を脇に押し付けるのに使われたかもしれないと言った。 [23] さらなる解釈は例えばB.テレサマリーナスカとハルツカオスモルスカ、骨ピンズハウジングハウスの塩腺 [30] 、または動物の防御の欠如を考慮して、動物により良い匂いを与える嗅覚組織の疑いがあるジョン・オストロムによって。 [最初に30]
これらの仮説のほとんどは、時間の経過とともに信用を失います。 [4] Zがありました。 B.シュノーケリング機能に必要な穴も、とにかくくちばしを備えた動物にほとんど感覚がない尾根の筋肉に近づくこともありません。空気吸収型として、これに利用できるスペースはあまりにも取るに足らないものであり、他のハドロサウルスもリーグでそれらをサポートすることなく大きな頭を持っていました。臭気神経と臭気細胞も骨蛇の外側にあり、後者は臭いの感覚とは何の関係もないことを示しています。 [24] 一方、これらの仮説では、骨コーンがこの形状を正確に持っていた理由や、他のランボオサウリンが頭にまったく異なる構造を着ていた理由を説明していません。代わりに、視覚的および聴覚機能は研究者に焦点を当てています。
視覚機能 [ 編集 | ソーステキストを編集します ]

大きなオブジェクトとして、骨円錐は視覚信号として使用できます。動物の目は非常に大きく、スケールリングを上げていました。これは、動物が一日活動的であり、視覚が非常に重要であることを示しています。おそらく、他のランボオサウリンで想定されているように、骨蛇の2つの視覚機能が与えられました。一方では、動物が他のハドロサウルスタイプの仲間の種であるように、際立った特徴としての機能を示しました( ランベオサウルス また コリトサウルス )、そして一方で、最近の鹿と同様に、性的識別(おそらくブライダル広告とライバルの印象のために)を区別することができます。ただし、これらの機能は、骨タップ内のチューブ長の複雑さを説明していないため、別の機能があったに違いありません。 [24]
共鳴体として機能します [ 編集 | ソーステキストを編集します ]
カール・ウィマンは、チューブの長さ(1931)のサウンド信号関数を提案した最初の人物であり、骨の鉛筆と曲がった角を比較しました。 [32] ホプソンとデイビッド・B・ワイハンペルは、1970年代と1980年代に再びこのアイデアを取り上げました。 [十] [33] [34] ホプソンは、ハドロサウルスンでの繊細な聴聞会への言及を見つけました:これは親relativeがどのようになったかです コリトサウルス 鼓膜のための大きな部屋と組み合わせて、敏感な中耳を示すあぶみ(ossicle)を発見しました。さらに、ラゲナはワニのように拡張されており、これはよく発達した内耳を示しています。 [十] Weihampelが予測しました Parasaurolophus Walker 骨タップの助けを借りて、48〜240 Hzの周波数を生成できたでしょう。ために Parasaurolophus cyrtocristatus 彼は75〜375 Hzを与えました。彼はまた、内耳の類似性とワニの類似性に基づいて、成人ハドロサウルデンは、若い動物を生産する可能性のある高周波数に敏感であると言いました。 [33]
よく保存された頭蓋骨 Parasaurolophus Tubicen コンピューターでモデル化され、3Dモデルに基づいて、動物によると可能性は霧のようなノイズを再構築しました。 [35] この種は、骨蛇の構造にさらに複雑さがあります Parasaurolophus Walker 、骨タップの主要部分は、約30 Hzのトーンを生成しました。 [36]
冷却のための機能 [ 編集 | ソーステキストを編集します ]
別の理論では、それは一連の温度調節であると述べています。 1978年に初めて提案されたこのアイデアは、1990年代以来再び議論されてきました。 P.E. Wheelerは、骨鉛筆が脳を冷却する手段であると最初に言いました。 [37] ほぼ同時に、骨鉛筆を通る温度調節のアイデアも、MaryańskaとOsmólskaによる論文に登場しました。この属について、サリバンとウィランソンによる研究の理論にさらに注意が払われました。 [38] 骨ピン内の大きな表面と血管は、証拠として名前が付けられています。 Lambeosaurinae Combsの機能についてのDavid Evans(2006)による議論の中で、彼はこのアイデアを支持しました。 [24]
最初の発見は、1920年にアルバータ州(カナダ)のレッドディアリバーにあるサンドクリークにあるトロント大学の遠征の一環として行われました。 [7] 岩層は現在、恐竜公園層として知られており、カンパニウムの化石が含まれています。コピー(ホロタイプ;コピー数ROM 768)は、頭蓋骨のある部分的な骨格で構成されており、膝関節と尾の大部分の骨の骨が欠落しています。

1922年、ウィリアム・パークスは新しい動物をと説明しました Parasaurolophus Walker 。公園は属と呼ばれました パラサウロロフス 「ほぼ紋章」と同じくらい。サイズ。 サウロス – トカゲ、 lophos – comb)それ以来意味します サウロロフス 考えは親しい親relativeでした。 arthepitheth ウォーカー 王立オンタリオ博物館の評議員会の議長であるバイロン・エドマンド・ウォーカーir。 [7] パラサウロロフス – しかし、アルバータ州からはまれなままでした [19] – だから、頭蓋骨のない恐竜公園のフォーメーションからはさらに3つの発見が知られているかもしれませんが、それもそうかもしれません パラサウロロフス 属する。 [39] 2007年、若い動物の脳の頭蓋骨(コピー数CMN 8502)が再検査され、このジャンルは起因しました。この発見は、おそらく恐竜のパーク層から、1917年にチャールズ・モルトラム・スターンバーグによって、1924年にギルモアによって収集されました。 Lambeosaurus sp。 説明された。 [13]
チャールズ・ヘイゼリウス・スターンバーグは、1921年にサンファンカントリー(ニューメキシコ)のやや若いカートランド層で断片的な頭蓋骨を発見しました。 1931年、カール・ウィマンは彼を第二の種類と説明しました、 Parasaurolophus Tubicen 。 [32] arthepitheth Tubicen ラテン語から派生したものであり、「トランペッター」を意味します。 [40] の2番目の、ほぼ完全な頭蓋骨 Parasaurolophus Tubicen (模範的な番号NMMNH P-25100)1995年にニューメキシコで発見されました。この頭蓋骨は、1999年にコンピューター断層撮影を使用して検査され、とりわけ、骨タップの機能に関する洞察を提供しました。 [8]
ジョン・オストロムは、1961年にニューメキシコの別の良い発見(コピー数FMNH P27393)を説明しました Parasaurolophus cyrtocristatus 。現在、自然史の野外博物館にあるコピーは、短い丸い骨蛇と後頭蓋骨格の大部分(頭蓋骨に接続されている残りの骨格)を備えた不完全な頭蓋骨で構成されています。尾の足、首、部分が欠けています。 [41] 種の名前はラテン語の言葉に導かれます cortured – 「短縮」と 紋付き – 「Cam -Bearing」。 [40] スケルトンは、フルーツランド層の上部に見られるか、上記のカートランド層で可能性が高いでしょう。 [8] 1979年、ユタ州でも種が実証されました。デビッドB.ワイハンペルとジェームズA.ジェンセンは、ガーフィールド郡カイパロウィッツ層の層から同様に短い骨蛇で不完全な頭蓋骨(コピー数BYU 2467)を説明しました。 [42] 1999年、ユタで作られた別の頭蓋骨が短く丸い骨円錐形で説明されました。 [8] 2013年に説明された少年の骨格は、1年前の若い動物でさえ小さな骨の櫛のみを示していません。 [43]
彼の壮大なホーンコーンで パラサウロロフス 多くの映画やテレビ番組の俳優。 1つの例は、子供向け映画シリーズ「私たちの時間の前に国」(時間の前の土地)で、「ダッキー」は公式に1つとして公式に パラサウロロフス 彼女がaであっても適用されます サウロロフス は。 [44] この恐竜は、ディズニーの「ファンタジア」の「ル・サクレ・ドゥ・プリント」のセクションのように、多くの映画の背景にも登場します。 [45] 3つの「ジュラシックパーク」映画すべて [46] ディズニーの「恐竜」 [47] 。さらに、あります パラサウロロフス 恐竜のおもちゃとしてしばしば何十年もの間、例えばB.「ルイマルクスアンドカンパニー」、「MPC」、「バタット」、「ハンコック」またはカーネギーコレクションから。
- ↑ グレゴリー・S・ポール: 恐竜へのプリンストンフィールドガイド。 プリンストン大学出版局、プリンストンニュージャージーu。 a。 2010、ISBN 978-0-691-13720-9、S。308、 オンライン 。
- ↑ a b c d そうです f ジョン・R・ホーナー、デビッド・B・ウェイシュンペル、キャサリン・A・フォースター: ハドロサウルス科。 In:David B. Weihampel、Peter Dodson、HalszkaOsmólska(編): 恐竜。 第2版。カリフォルニア大学出版局、バークレーCA u。 a。 2004、ISBN 0-520-24209-2、S。438–463。
- ↑ リチャード・S・ラル、ネルダ・E・ライト: 北米のハドロサウルスの恐竜 (= アメリカ地質学会。特別な論文。 bd。 40、 ISSN 0072-1077 )。アメリカ地質学会、ニューヨークNY 1942、S。229。
- ↑ a b c d そうです ドナルドF.大gut: パラサウロロフス。 In:Donald F. Glut: 恐竜。百科事典。 McFarland&Co、Jefferson NC u。 a。 1997、ISBN 0-89950-917-7、S。678–684。
- ↑ a b c リチャード・S・ラル、ネルダ・E・ライト: 北米のハドロサウルスの恐竜 (= アメリカ地質学会。特別な論文。 bd。 40、 ISSN 0072-1077 )。アメリカ地質学会、ニューヨークNY 1942、S。209–213。
- ↑ マイケル・K・ブレット・サーマン、ジョナサン・R・ワグナー: カンパニアンおよびマースチャンティアン北米のハドロサウルス群の虫垂解剖学。 In:Kenneth Carpenter(hrsg。): 角とくちばし。セラトプシアとornithopodの恐竜。 インディアナ大学出版局、2007年、ブルーミントン、ISBN 978-0-253-34817-3、S。135–169。
- ↑ a b c d そうです ウィリアム・A・パークス: Parasaurolophus Walkeri。 Crested Trachodont Dinosaurの新しい属と種 (= トロント大学研究。地質学シリーズ。 bd。 13、 ISSN 0372-4913 )。トロント大学 – 大学図書館、トロント1922年、 デジタル化 。
- ↑ a b c d そうです ロバート・M・サリバン、トーマス・E・ウィリアムソン: ニューメキシコ州のカートランド層からのパラサウルスロフス(ディニノサウリア:ハドロサウルス科)の新しい頭蓋骨と属の改訂 (= ニューメキシコ自然史と科学博物館。速報。 15、 ISSN 1524-4156 )。ニューメキシコ自然史博物館、アルバカーキNM 1999、 デジタル化 。
- ↑ ダグラス・パーマー(編): 恐竜と先史時代の動物。イラスト付き百科事典。 ケーネマン、ケルン2000、ISBN 3-8290-6113-7、p。153。
- ↑ a b c d ジェームズ・A・ホプソン: ハドロサウルの恐竜における頭蓋展示構造の進化。 の: 古生物学。 bd。 1、nr。 1、1975、 ISSN 0094-8373 、S。21–43。
- ↑ チャールズ・W・ギルモア: ステファノサウルス属で、パークスのランベオサウルスランベイのタイプ標本の説明があります。 の: カナダ、鉱山省、地質調査。速報。 bd。 38、nr。 43、1924、 ZDB-ID 429582-1 、S。29–48。
- ↑ Pascal Godefroit、Shuqin Zan、Liyong Jin: Charonosaurus Jiaynensis N.G.G.、N.Sp.、中国北部の故MaastrictianのLambeosaurine Dinnos。 の: 科学アカデミーの報告。 シリーズ2、ブックレットA: 地球と惑星科学。 bd。 330、nr。 12、2000、 ISSN 0764-4450 、S。875–882、2: 10.1016/s1251-8050(00)00214-7 。
- ↑ a b c d デビッド・C・エヴァンス、ロバート・R・レイス、ケビン・デュプイス: アルバータ州の恐竜州公園の脳内ケース(オルニチシア:ハドロサウルス科)の脳ケース。 の: Journal of Vertebrate古生物学。 bd。 27、nr。 3、2007、 ISSN 0272-4634 、S。642–650、2: 10.1671/0272-4634(2007)27 [642:AJPOHB] 2.0.co; 2 。
- ↑ デイビッド・B・ウェイシュンペル、ジャック・R・ホーナー: ハドロサウルス科。 In:David B. Weihampel、Peter Dodson、HalszkaOsmólska(編): 恐竜。 カリフォルニア大学出版局、バークレーCA u。 a。 1990、ISBN 0-520-06726-6、S。534–561。
- ↑ a b c d デビッド・C・エヴァンス、ロバート・R・レイズ: アルバータ州恐竜公園層からの紋付きのハドロサウルスの恐竜(ornithischia)であるランベオサウルスマグニックリスタスの解剖学と関係。 の: Journal of Vertebrate古生物学。 bd。27、no。 2、2007、S。373–393、2: 10.1671/0272-4634(2007)27 [373:Aarolm] 2.0.co; 2 。
- ↑ a b c d トーマス・E・ウィリアムソン: ニューメキシコ州サンファン盆地からのハドロサウルス科(ディノサウリア、オリチシア)のレビュー。 In:Spencer G. Lucas、Andrew B. Heckert(hrsg。): ニューメキシコの恐竜 (= ニューメキシコ自然史と科学博物館。速報。 17)。ニューメキシコ自然史博物館、アルバカーキNM 2000、S。191–213、 オンライン 。
- ↑ a b デビッドA.エバース: 地質。 In:Philip J. Currie、Eva B. Koppelhus(編): 恐竜の地方公園。壮大な古代の生態系が明らかになりました。 インディアナ大学出版局、ブルーミントンのU。 a。 2005、ISBN 0-253-34595-2、S。54–82。
- ↑ a b David B. Weishampel、Paul M. Barrett、Rodolfo Coria、Jean Le Loeuff、Xing Xing Xing Xijin Zhao、Ashok Sahni、Elizabeth Gomani、Christopher R. Noto: 恐竜の分布。 In:David B. Weihampel、Peter Dodson、HalszkaOsmólska(編): 恐竜。 第2版。カリフォルニア大学出版局、バークレーCA u。 a。 2004、ISBN 0-520-24209-2、S。517–683、Hier S. 517–606。
- ↑ a b マイケル・J・ライアン、デビッド・C・エヴァンス: Ornithischian Dinosaurs。 In:Philip J. Currie、Eva B. Koppelhus(編): 恐竜の地方公園。壮大な古代の生態系が明らかになりました。 インディアナ大学出版局、ブルーミントンのU。 a。 2005、ISBN 0-253-34595-2、S。312–348。
- ↑ デニス・R・ブラマン、エヴァ・B・コッペルフス(編): カンパニアのパリノモルフ。 In:Philip J. Currie、Eva B. Koppelhus(編): 恐竜の地方公園。壮大な古代の生態系が明らかになりました。 インディアナ大学出版局、ブルーミントンのU。 a。 2005、ISBN 0-253-34595-2、S。101–130。
- ↑ デールA.ラッセル: 時間内のオデッセイ。北米の恐竜。 ノースワードプレスu。 a。、Minocqua wi u。 a。 1989、ISBN 1-55971-038-1、S。160–164。
- ↑ ロバート・T・バッカー: 恐竜の遺伝。新しい理論は恐竜の謎とその絶滅を解き放ちます。 ウィリアム・モロー、ニューヨークニューヨーク1986、ISBN 0-688-04287-2、S。194。
- ↑ a b c デビッド・B・ノーマン: ハドロサウルスII。 In:David B. Norman: 恐竜のイラスト入り百科事典。恐竜の王国における生活に対する独創的で説得力のある洞察。 Crescent Books、ニューヨークNY 1985、 ISBN 0-517-468905 、S。122–127。
- ↑ a b c d デビッド・C・エヴァンス: ランベオサウリンの恐竜における鼻腔ホモロジーと頭蓋紋章機能。 の: 古生物学。 bd。32、no。 1、2006、S。109–125、2: 10.1666/0094-8373(2006)032 [0109:NCHACC] 2.0.co; 2 。
- ↑ アルフレッド・シャーウッド・ロマー: 脊椎動物の古生物学。 シカゴ大学出版局、シカゴイリノイ州1933年、S。491。
- ↑ マーティン・ウィルファース: 回復力のある恐竜。 の: オリオン。自然と技術のためのイラスト誌。 bd。 2、1947、 ZDB-ID 207436-9 、S。525–532。
- ↑ チャールズ・M・スターンバーグ: 白亜紀後期の腹川シリーズのフード付きハドロサウル (= カナダ国立博物館。地質シリーズ。速報。 52、 ZDB-ID 429579-1 = カナダ国立博物館。速報。 77). Patenaude, Ottawa 1935.
- ↑ エドウィンH.コルバート: 恐竜の本。支配爬虫類とその親relative (= 人と自然の出版物。ハンドブック。 nr。 14)。アメリカ自然史博物館、1945年ニューヨークニューヨーク、 デジタル化 。
- ↑ オセニオ・アベル: 新しい恐竜はOberkreide Canadasで見つけます。 の: 自然科学。 bd。12、no。 36、1924、S。709–716、2: 10.1007 / BF01504818 。
- ↑ TeresaMaryańska、HalszkaOsmólska: ハドロサウルスの頭蓋解剖学の側面。 の: レダイア。 bd。 12、nr。 3、1979、 ISSN 0024-1164 、S。265–273、2: 10.1111/j.1502-3931.1979.tb01006.x 。
- ↑ ジョン・H・オストロム: ハドロサウルの恐竜の頭蓋紋章。 の: 郵便で。 bd。 62、1962、 ISSN 0079-4295 、S。1–29。
- ↑ a b カール・ウィマン: Parasaurolophus tubicenn。sp。ニューメキシコのチョークから (= ウプサラのActa Regiae Society。 シリーズ4、Vol。7、No。5、 ZDB-ID 210356-4 )。 Almqvist&Wiksell、Uppsala 1931。
- ↑ a b David B. Weihampel: Lambeosaurine Dinosaurs(爬虫類:Ornithischia)における潜在的な発声の音響分析。 の: 古生物学。 bd。 7、nr。 2、1981、S。252–261。
- ↑ David B. Weihampel: Lambeosaurine Hadrosaurids(爬虫類; Ornithischia)の鼻腔;比較解剖学と相同体。 の: Journal of Paleontology。 bd。 55、nr。 5、1981、S。1046–1057。
- ↑ サンディア国立研究所: 科学者はデジタル古生物学を使用して、パラサウロロフス恐竜の声を生み出します。 1997年12月5日、アーカイブ オリジナル 午前 17. 2014年8月 ; 2014年10月24日に取得 。
- ↑ カール・F・ディーゲルト、トーマス・E・ウィリアムソン: Lambeosaurine Hadrosaur Parasaurolophus Tubicenのデジタル音響モデル。 の: Journal of Vertebrate古生物学。 Vol。18、No。3=の補足 第58回年次総会、脊椎動物古生物学協会、スノーバードスキーアンドサマーリゾートスノーバード、ユタ州スノーバード、1998年9月30日 – 10月3日 、1998、S。38a。
- ↑ P. E.ウィーラー: 大きな恐竜の精巧なCNS冷却構造。 の: 自然。 bd。275、no。 5679、1978、S。441–443、2: 10.1038/275441a0 。
- ↑ ロバート・M・サリバン、トーマス・E・ウィリアムソン: の新しい頭蓋骨 パラサウロロフス ニューメキシコ州からの(長いクレストされた形式):外部および内部(CTスキャン)機能とその機能的意味。 の: Journal of Vertebrate古生物学。 Vol。16、No。3=の補足 1996年10月16〜19日、ニューヨーク州ニューヨーク州自然史博物館、アメリカ自然史博物館、脊椎動物古生物学会、第56回年次総会の要約 、1996、pp。68a。
- ↑ フィリップ・J・カリー、デール・A・ラッセル: 恐竜の地理的および層序分布の遺物。 In:Philip J. Currie、Eva B. Koppelhus(編): 恐竜の地方公園。壮大な古代の生態系が明らかになりました。 インディアナ大学出版局、ブルーミントンのU。 a。 2005、ISBN 0-253-34595-2、S。553。
- ↑ a b デビッド・P・シンプソン: カッセルのラテン – 英語、英語 – ラテン語辞書。 第5版。カッセルu。 a。、ロンドンu。 a。 1979、S。883。
- ↑ ジョン・ハロルド・オストロム: ニューメキシコ州の白亜紀からのハドロサウルスの新しい恐竜の新種。 の: Journal of Paleontology。 bd。 35、nr。 3、1961、S。575–577。
- ↑ デビッド・B・ウェアハンプル、ジェームズ・A・ジェンセン: ユタ州のパラサウロロフス(爬虫類;ハドロサウルス科)。 の: Journal of Paleontology。 bd。 53、nr。 6、1979、S。1422–1427。
- ↑ アンドリュー・A・ファーク、デレク・J・チョホック、アニサ・ヘレロ、ブランドン・スコリエリ、サラ・ワーニング: チューブが押し付けられた恐竜のパラサウロロフス(ハドロサウルス科)の個体発生とハドロサウルスの不整合。 の: Peerj。 、22。2013年10月、1:E182 doi: 10.7717/peerj.182 。
- ↑ 「私たちの時間の前の国で」DVD ( 記念 2009年2月17日から インターネットアーカイブ ))
- ↑ ファンタジー(1940)。 の: 映画の間違い。 2014年10月24日に取得 。
- ↑ パラサウロロフス。 の: 公園は尋ねた。 2014年10月24日に取得 。
- ↑ ディズニー・恐竜はデビッド・クレンツにインタビューします。 の: デイブ・ホーンのオーキサウルスの黙想。 2014年10月24日に取得 。
Recent Comments