横方向阻害 – ウィキペディア
いつ 横方向の阻害 (また 環境の阻害 、 ラテーレ阻害 、 横方向の阻害 また ラテラリンヒッション 神経細胞の回路原理と呼ばれる神経生物学では、神経生物学において、活性神経細胞が隣接細胞の活性を阻害します。
この一般的な神経生理学的原理は、中枢神経系全体で発生します – 最もよく研究された例は、箸の急速なケーブルを阻害する日光による円錐の興奮です。 [初め] [2]
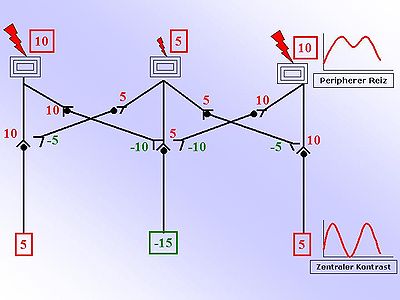
右側の図面は、この原則の理解を簡素化します。たとえば、皮膚の表面に3つの受容体を見ることができます。 2つの外側のものは、内側の受容体よりも興奮しています。隣接する数字は、強度に関する情報を提供する相対ユニットです。数が大きいほど、興奮が大きくなります。正の赤い数字は刺激的で、負で、緑の数字は抑制性です。興奮の強さは現在、活動電位周波数に変換され、転送されます。示されている表示された中間スキームは、刺激的な受信シグナル(求心性)を抑制性(抑制性)継続信号に変換する抑制性内ロンを示しています。 スイッチ 興奮から抑制まで、異なる神経伝達物質を使用して行われます(阻害:GABA、グリシン、アドレナリン) [3] 。この抑制性の内部ニューロンは、二次伝播により、SO -Called 2番目のニューロンへの脆弱性が弱まります。この弱体化は、インターネットの興奮に比例します(抑制:図のネガティブ、グリーン数)。2番目のニューロンに対する刺激的で抑制的な影響を単純に合計することにより、コントラストの強化があります。これは、隣接する図によってグラフィカルに表示されます。
それは、ガングリエンゼルノードにあるものや網膜の上層の目にある複雑な絡み合った回路で発生します。 [4]
網膜では、抑制性内部ニューロンは水平細胞と呼ばれ、光受容体の側面に切り替えるのに役立ちます(箸と円錐)。 [5] ただし、画像のコントラストと動きの迅速な検出を強化するために、網膜には切り替えのための他の複雑なオプションがあります。
横方向の阻害の結果は、何よりもコントラスト補強、したがって形状制限の発達であり、空間方向の基礎として機能します。
横方向の阻害の概念は、発達生物学における調節ルートも説明しています。たとえば、緊急信号経路によって伝えられます。マルチセラーの開発は複雑なプロセスであるため、増殖と専門化の正確な制御が必要です。横方向の阻害は重要な役割を果たすと想定されています。 [6] 横方向の阻害の場合、通常、細胞のさらなる発達に関するバイナリ決定が行われます。 [7]
ここでは、すべての細胞は、最初は、細胞が発現するすべての周囲の細胞よりも確率的プロセスとより多くのデルタに関連する非対称性を持つまで、同じ「タイプ」です。この非対称性は次の肯定的なフィードバックを強化し、他の細胞では、デルタが吸入され、より多くの緊急事態が表されます。したがって、これらの細胞は他の「タイプ」の細胞になります。 [8]
たとえば、このプロセスは、有毛細胞の正確な構造のトレーニングと耳の細胞を支える役割を果たします。鶏の場合、Notchが最初にSo -Calcaled Colsensory細胞の形成を開始し、横方向の阻害が最終的な毛細胞の量と分布を調節することが実証されました。 [9]
人工ニューラルネットワークのインスピレーションとしての横方向の阻害(機械学習) [ 編集 | ソーステキストを編集します ]
いわゆる「畳み込みニューラルネットワーク」(CNNS)では、横方向の阻害のプロセスは、層内の最大プーリングステップで最も活性なニューロンに応じてニューロンを選択することによりシミュレートされます。 [十] 、隣接する非アクティブなニューロンは、ネットワークのさらなる計算ステップのためにオフになります。 [11]
- ↑ Ashish Bakshi、Kuntal Ghosh: 視覚の知覚された明るさを計算するための注意とフィードバックのニューラルモデル 。の: 神経計算のハンドブック 。 Elsevier、2017、ISBN 978-0-12-811318-9、 S. 487–513 、doi: 10.1016/b978-0-12-811318-900026-0 ( Elsevier.com [2019年10月23日にアクセス])。
- ↑ フリードリッヒ・ゼトラー、マティ・ジェルヴィレト: 昆虫の目の横方向の阻害 。の: 比較生理学のための雑誌 。 バンド 76 、 いいえ。 3 、1972年、ISSN 0340-7594 、 S. 233–244 、doi: 10.1007/BF00303230 ( springer.com [2019年10月25日にアクセス])。
- ↑ リチャード・H・クレイマー、クリストファー・M・ダベンポート: 脊椎動物網膜の横方向阻害:欠落している神経伝達物質の場合 。の: PLOS生物学 。 バンド 13 、 いいえ。 12番目 、2015年12月10日、ISSN 1545-7885 、 S. E1002322 、doi: 10.1371/journal.pbio.1002322 、 PMID 26656622 、 PMC 4675548 (無料の全文) – ( plos.org [2019年10月23日にアクセス])。
- ↑ キャメロンH.G.ライト、スティーブンF.バレット: 生体模倣視覚センサー 。の: エンジニアリングバイオミミクリー 。 Elsevier、2013、ISBN 978-0-12-415995-2、 S. 1–36 、doi: 10.1016/b978-0-12-415995-2.00001-5 ( Elsevier.com [2019年10月23日にアクセス])。
- ↑ WebVision:網膜と視覚システムの組織 。ユタ大学健康科学センター、ソルトレイクシティ(UT)1995、カップ。 「網膜の単純な解剖学」、 PMID 21413389 ( NIH.GOV [2019年10月23日にアクセス])。
- ↑ ブルース・アペル、リー・アン・ギバン、ジュディス・S・アイゼン: [タイトルは見つかりません] 。の: BMC発達生物学 。 バンド 初め 、 いいえ。 初め 、2001年、 S. 13 、doi: 10.1186/1471-213X-13 、 PMID 11495630 、 PMC 37243 (無料の全文) – ( BiomedCentral.com [2019年10月24日にアクセス])。
- ↑ Makoto Sato, Tetsuo Yasugi, Yoshiaki Minami, Takashi Miura, Masaharu Nagayama: Notchを介した横方向阻害は、EGFを介した反応拡散と組み合わせると、発生波の伝播を調節します 。の: 国立科学アカデミーの議事録 。 バンド 113 、 いいえ。 35 、30。2016年8月、ISSN 0027-8424 、 S. E5153 -E5162 、doi: 10.1073/pnas.1602739113 、 PMID 27535937 、 PMC 5024646 (無料の全文) – ( pnas.org [2019年10月24日にアクセス])。
- ↑ N. Perrimon、C。Pitsouli、B.-Z.シロ: 細胞の運命と胚のパターニングを制御するシグナル伝達メカニズム 。の: 生物学におけるコールドスプリングハーバーの視点 。 バンド 4 、 いいえ。 8 、1。2012年8月、ISSN 1943-0264 、 S. A005975 -A005975 、doi: 10.1101/cshperspect.A005975 、 PMID 22855721 、 PMC 3405863 (無料の全文) – ( cshlp.org [2019年10月24日にアクセス])。
- ↑ N. Daudet: ひよこの内耳の発達におけるノッチ活動の2つの対照的な役割:補綴パッチの仕様と毛細胞分化の横方向の阻害 。の: 発達 。 バンド 132 、 いいえ。 3 、5。2005年1月、ISSN 0950-1991 、 S. 541–551 、doi: 10.1242/dev.01589 ( Biologists.org [2019年10月25日にアクセス])。
- ↑ Cao、C.、Huang、Y.、Wang、Z.、Wang、L.、L.、N。、&Tan、T。: 視覚的注意と顕著性検出のための横方向の抑制に触発された畳み込みニューラルネットワーク 。の: 人工知能に関する32秒のAAAI会議 。 2018年4月。
- ↑ ブルーノ・ホセ・トーレス・フェルナンデス、ジョージ・D・C・カヴァルカンティ、ツァン・レン: 画像分類のための横方向の阻害錐体ニューラルネットワーク 。の: サイバネティックスに関するIEEEトランザクション 。 バンド 43 、 いいえ。 6 、2013年12月、ISSN 2168-2267 、 S. 2082–2092 、doi: 10.1109/tcyb.2013.2240295 ( IEEE.org [2019年11月1日にアクセス])。
Recent Comments