Asiatischer Esel – ウィキペディア
アジアのロバ ( 馬のhemionus )馬の属からの野生のロバです( エクウス )馬の家族(equidae)内。彼はアフリカのロバです( 馬のロバ ) – 家のトランク形態と同様ですが、多くの馬のような特性もあります。このため、彼もそうなるでしょう 半分 また PFERDEESEL 専用。また、「Khur」や「Kulan」などの多くの地域名でも知られています(Systematicsを参照)。アジアのロバは、西部から中央および北アジアまでの乾燥した半砂漠や草原のような風景に住んでおり、主に硬い草の食べ物を食べています。アジアのロバの社会構造は非常に複雑であると考えられていますが、種はまだ詳細に調査されていません。全体として、株式は絶滅危ed種と見なされます。
習慣 [ 編集 | ソーステキストを編集します ]

頭部胴体の長さは200 cmを超え、尾の長さは40 cm、肩の高さは97〜138 cm、200〜260 kgの重量で、アジアのロバは平均的な家よりわずかに大きくなっています。セクシュアモルフィズムはわずかに顕著です。一般に、馬のタイプは長い脚でスリムです。ひづめはアフリカのロバよりも広い( 馬のロバ )。すべてのシマウマやロバと同様に、特徴 栗 、茶色のカルスのような調査、前足のみ、野生の馬とは対照的に、後肢にも現れます。耳も本物のロバの耳よりも長いです。さらに、テールタッセルとたてがみはあまり顕著ではなく、食事は依然として典型的です。上部は、短い髪の夏の毛皮で灰色で、黄色から黄土色の毛、または赤茶色です。下側、首と脚は白または部分的にクリーム色です。個々の亜種は、毛皮の色の強度が異なります。ただし、ボディページの色の移行は、あらゆる形態の鎌の月のようなコースによって特徴付けられます。これは側面に部分的に引き上げられますが、野生の馬のこの移行のm字型とは大きく異なります。長い髪の冬の毛皮は一般に暗い設計されています。夏には70〜80、冬には70〜90 mmです。このバックストリップは、多くの場合、軽い毛皮の部分で囲まれています。アフリカのロバに典型的な水平脚ストリップパターンは、アジアのロバではめったに起こりません。 [初め] [2] [3]
頭蓋骨と歯列の特性 [ 編集 | ソーステキストを編集します ]

アジアのロバの頭蓋骨は長さ42〜52 cmで、キアンの頭蓋骨です( エクウス・キアン )典型的な短い鼻と非常によく似ています。最前線は比較的まっすぐに動作し、ボタンは狭く垂直で、頸椎(顆)の基部表面を越えて引っ張られません。鼻骨の長さは最大22 cmで、前面にわずかな曲率があります。鼻の骨と中間顎骨の間の鼻の内部と上顎の内側は比較的大きいです。アイソケットは頭蓋骨では非常に低いことがありますが、常に最後の臼歯の後ろにあります。 [初め] [4]
大きな下顎の長さは最大42 cmに達し、顎骨の幅は最大5 cmです。特に顎関節は非常に強いです。成体動物の歯の式は次のとおりです。
。切歯は非常に幅が広く、顎の斜面ではキアンよりも前向きです( エクウス・キアン )、角の歯は通常小さいです。背中に利用できる拡張腫は、幅8.6 cmまでのものにすることができます。臼歯は臼歯と同様に構造化されています。つまり、明らかに臼歯化され、時には最初の臼歯は初歩的です(オオカミの歯)。一般に、臼歯には、多くの歯科用セメントと巻き歯の溶融ストリップが高くなります。エナメル質は、下臼歯の購入スペースで2つの突起の間を走ります メタコニド と メタスチリド 背面(舌側)の領域では部分的にV字型ですが、時にはより明確です。 [初め]
 。切歯は非常に幅が広く、顎の斜面ではキアンよりも前向きです( エクウス・キアン )、角の歯は通常小さいです。背中に利用できる拡張腫は、幅8.6 cmまでのものにすることができます。臼歯は臼歯と同様に構造化されています。つまり、明らかに臼歯化され、時には最初の臼歯は初歩的です(オオカミの歯)。一般に、臼歯には、多くの歯科用セメントと巻き歯の溶融ストリップが高くなります。エナメル質は、下臼歯の購入スペースで2つの突起の間を走ります メタコニド と メタスチリド 背面(舌側)の領域では部分的にV字型ですが、時にはより明確です。 [初め]
。切歯は非常に幅が広く、顎の斜面ではキアンよりも前向きです( エクウス・キアン )、角の歯は通常小さいです。背中に利用できる拡張腫は、幅8.6 cmまでのものにすることができます。臼歯は臼歯と同様に構造化されています。つまり、明らかに臼歯化され、時には最初の臼歯は初歩的です(オオカミの歯)。一般に、臼歯には、多くの歯科用セメントと巻き歯の溶融ストリップが高くなります。エナメル質は、下臼歯の購入スペースで2つの突起の間を走ります メタコニド と メタスチリド 背面(舌側)の領域では部分的にV字型ですが、時にはより明確です。 [初め] 感覚のパフォーマンスと音の表現 [ 編集 | ソーステキストを編集します ]
他の馬と同様に、アジアのロバの主なコミュニケーションは、排泄された分泌物についての嗅覚ですが、視覚と良い聴覚も良い聴覚を持っています。このようにして、彼らは通常、潜在的な危険まで2 kmの距離で飛行します。 [5] [6] これまでのところ、彼らの健全な表現に関する調査はほとんどありませんでした。
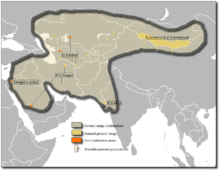
以前(オリーブ)と今日(黄色とオレンジ)
アジアのロバの流通エリアは、西アジアの地中海地域からトランスカカシア、中央アジアまで、南アジアの西部地域と北アジアの南部地域までの範囲です。しかし、今日では、流通エリアは強く粉砕されているため、アジアのロバはモンゴル、トルクメニスタン、カザフスタン、インド、イラン、ウクライナ、サウジアラビアの個々の地域に限定されています。 Dschiggetaiは主にゴビ砂漠とKhurのKhurのKachchhの走りに住んでいます。クランは、バッドキッツ自然保護区の大勢の人口とアンダサシスキ自然保護区で見つけることができますが、オナガーはトゥーラン保護エリアとバーラム・エ・ゴアにまだ見られます。 [3]
アジアのロンキーの生息地は、乾燥した半砂利、草原、山の階段、部分的に砂漠です。これらの極端な生息地は、海抜2000 mまでに達する可能性がありますが、夏の高温(最大40°C)、冬には低い(最大-35°C)、時には100 mm未満の低降水量が特徴です。さらに、エリアは薄い植生の天井によって特徴付けられます。 [3] [7] その流通エリアの西部では、アジアのロバは実際のロバと一緒に現れていましたが、より深いエリアを好みました。今日、両方のタイプが荒野で根絶されています。 [8] 東では、プリヴァルスキー馬の分布エリアと部分的に重複しています( Equus przewalskii )。 [6]
領土行動 [ 編集 | ソーステキストを編集します ]

アジアのロバの社会構造は非常に異なっています。ゴビ砂漠とクラン中央アジアのdschiggetaiによると、北部の個体群は、種牡馬、いくつかの牝馬と子馬、大きなアクションルームを備えた群れの形成である傾向があります( ホームレンジ )。また、ここには大規模なハイキングがあります。これには、4,500〜40,000km²のエリアが含まれ、夏のハイキングは冬よりも制限されています。場合によっては、最大450人の個人の大規模なグループ関連も形成されます。これは通常、良い食品の場所や水ポイントでのみ行われます。これらの大規模な関連付けは1日以内に溶けているため、個々の群れのランキングに加えて、上位の階層はないようです。冬にしばしば形成される若い雄動物の「独身グループ」もあります。そのような生き方は、野生の馬と草原からのものです( Equus Quagga )そしてBergzebra( エクウスゼブラ )既知。 [9] [6] [十] しかし、一時的な領土の形成も時々あり、それは積極的に防御されます。 [11]
KhurとOnagerによると、南部の個体群は、純粋に領土のライフスタイルになる傾向があり、それにより地域が部分的に重複しています。支配的な種牡馬は、サイズが9km²に達するが、大幅に大きくなる可能性があるエリアを維持します。これらの領土には、食料と休息エリア、恒久的または定期的な水源が含まれています。ただし、水は通常、領土の端にあり、中央にはありません。頻繁にコミットされる経路には、糞と尿がマークされており、同じマーキングポイントが使用されています。子馬の症状は、最大20km²の領域を放牧する小さなグループに形成されることがあり、他のグループや支配的な種馬の領域と重複しています。同様の動作はGrevyzebraからです( ホースグレヴィ )そしてアフリカのロバから知られています。 [12番目] [5] [13]
アジアのロバのさまざまな社会的行動は完全に明らかにされていません。気候因子または狩猟圧力は、獲物の側で役割を果たす可能性があります。 KhursやOnagersの配電室には、より大きな捕食者がほとんどいないため、領土の動物よりも元々のライフスタイルを保持しています。一方、KulanとJiggetaiは、より安定したグループが生存の可能性が高いため、群れの形成を刺激した可能性のあるオオカミの狩猟圧力にさらされています。ここで、種牡馬はそのグループを積極的に擁護します。 [9]
栄養 [ 編集 | ソーステキストを編集します ]
アジアのロバの食事による詳細な研究はわずかしかありません。一般に、高溝の臼歯と歯科用の大部分を備えた硬い小石に適応しています。 放牧 )。ゴビの個体群は、居住したステップの生息地(Stipa steppe)で広範囲に発生する羽の草を好みますが、尾根もぶつかります アチナサラム – グラス、リード、ビン。証明されています アルテミシア 、 アナバシス 、塩ハーブ、サクサール、エンドウ豆の茂みがアジアのロバの食物のスペクトルにあります。 [9] [14] さらに、Lauchも地域に来ます、 Zygophyllum 食物植物として存在するタマリスク植物。 [15] 南部の個体群の場合、フィンガーチッシュ、倒れた種子の草、愛の草、そして ジカンチウム – 草は食物源として見つかりました。 [16] ただし、乾燥時には草の成長が枯れた場合にのみ枯れている場合、アジアのロバは、特に存在しない限り、追加の木質植物を消費します。彼はひづめを使って木材を壊して、植物のより多くの水を含む領域を手に入れます。種子の吸収も観察されました。 [3]
流通エリアの乾燥気候条件のため、アジアのロバは絶えず利用可能な水を必要とします。これは10〜15 kmを超えてはなりませんが、水域への長さ30 kmのハイキングも観察されています。アジアのロバはまた、水を求めて最大60 cmの深さを掘っています。冬には、動物も雪を消費します。 [9] [3]
再生 [ 編集 | ソーステキストを編集します ]

アジアのロバの繁殖挙動もほとんど検査されておらず、野生ではめったに観察されませんでした。アジアのロバは平均して2年で性的に成熟しており、最初のペアリングは通常3〜4年で行われます。ペアリングは4月から9月の間に行われますが、6月と7月には蓄積があります。妊娠期間は約11ヶ月続き、それに応じて1つの子馬が生まれ、出生にはわずか10分しかかかりません。これは15〜20分以内に立つことができ、最新の1時間半後に牛乳を吸い始めます。飼育中、子馬と母親の動物は近くにとどまり、自分の年配の若い動物を含む他の動物が母なる動物に置かれます。時折、領土的に生きている人口の種牡馬は、若い動物を追い出し、雌馬を覆おうとします。野生のアジアのロバは14歳に達し、飼育下では最大26歳になる可能性があります。 [9] [17] [18]
他の動物種との相互作用 [ 編集 | ソーステキストを編集します ]
最大の捕食者はオオカミです。研究では、獲物の割合の23%がアジアのロバに割り当てられたことが示されました。馬のタイプは捕食者に対して積極的に自らを守りますが、逃げるアジアのロバも非常に速く、ギャロップで70 km/hの速度に達することができます。時折、甲状腺腫のガズ細胞がアジアのロバのグループの近くに現れます。 [9] [6]
寄生虫 [ 編集 | ソーステキストを編集します ]
最も一般的な内側の寄生虫には、主に糸虫が含まれます。 Trichostrongylus 、 パラスカリス 、馬の宮殿ワーム、肺虫、ドワーフ糸ワーム。さらに、サクシジアへの侵入を含む、サナダムシも発生します。 アイメリア 知られている。 Przewalski馬と家の馬で相互感染が発生する可能性があることは注目に値します。 [19]
|
属の内部システム エクウス Nach Vilstrup et al。 2013年 [20]
|
アジアのロバは属の代表として属します エクウス 今日の現代の馬。下臼歯の形態は非常に多様ですが、ステノニンまたは非カバリンの馬のグループにはまだ配置されています。 [21] 2009年のDNA分析は、キアンのアジアのロバの次の親relative( エクウス・キアン ) は。 [22] ヨーロッパの野生のロバ( Hydruntinus )それは、ヨーロッパとアジアの更新世中期および上部で広まっており、完新世でしか消えてしまった可能性があります。これは今日のアジアのロバよりもスリムで大きく、肩の高さは160 cmに達しました。 [2] もともと、一部の専門家はこのタイプの馬をアフリカのロバの近くに置いていましたが、解剖学的研究はアジアのロバとかなり緊密な関係を示しました。 [23] これは遺伝子分析によって確認できます。 [24] アジアのロバとキアンとヨーロッパの野生のロバの両方がステノニンの馬の中にあります ヘミオニン アジアのロバの学名にあるものを説明します 馬のhemionus 彼らの密接な関係を参照し、表現します。おそらく密接に関連する馬の種は、さまざまなロバとシマウマを表しています。 エクウス 。 [25] [22] [26] 学名 馬のhemionus 1775年にピーターサイモンパラスによって紹介された最初の説明は、モンゴル北東部の個人に基づいて行われました。 [27] [9]
現在、アジアのロバには6つの亜種がありますが、そのうち2つは絶滅しています。 [28]
- アナトリアの半分( E. h。 anatoliensis Haltenorth&Bridle 、1956);アナトリア、絶滅
- モンゴルの半分、地元のdschiggetai( E. h。 hemionus パラス 、1775);モンゴル
- シリアの半分、地元のアチダリ( E. h。裾 I. Goeffroy 、1855);シリア、メソポタミア、アラビア、1927年頃に時代遅れ
- インドの半分、地元のクル( E. h。 Khur レッスン 、1827);イラン南部、パキスタン、インド北西部
- トランスキャスピックまたはトルクメニッシュハルベッセル、地元のクラン( E. H.ハナン ( Groves&Mazák 、1967));カザフスタン、トルクメニスタン
- ペルシャの半分またはオナガー、地元のgur-khar( E. h。 onager ロッダ 、1785);北イラン
もともとは、亜種への分裂について多くの意見の相違がありました。多くの古い作品では、7〜8種類のアジアのロバがまだ区別されています。これは通常、今日の亜種として分類されています。例外は、今日ではそれ自身の種類を表しているKiangです。 [初め] Colin Peter GrovesとPeter Grubb、The Khur(Khur)が発表した2011年の蹄動物の改訂で Equus Khur )そしてachdari( 馬の裾 )独立した種として認識されています。 [29]

異なる亜種は、分布エリアの南西から北東へのサイズの増加によって特徴付けられます。南西部のアチダリは肩の高さが100 cmしか伸びていませんでしたが、ジゲゲタイは北東部で127〜138 cmです。その後、クランは高さ108〜116 cmになり、OnagerとKhur 112〜120 cmの肩の高さが与えられます。 [初め] 一部の当局の見解では、OnagerとTrangniesは同じ亜種です。馬の系統発生に関する現在の分子遺伝学的研究によれば、両方の集団は互いに明確に区別される可能性があります。毛皮の色が異なるため、さらなる亜種がジゲトイから分割されることがあります(ゴビハルベセル) horse hemionus黄色 )、、 [初め] しかし、他の専門家によると、これらの他の専門家は個々の集団で非常に多様です。 [9] 2009年のDNA検査では、オナガーが山のゼブラに近いため、すべての亜種が単系統由来であることが確認されていません。 [22] 同じ年の別の研究は、アジアのロバのすべての亜種の密接な関係を示しています。 [25]
|
種の内部システム 馬のhemionus ベネットの後、それは古い。 2017年 [30]
|
2017年の亜領土および最近の材料のより包括的な遺伝分析は、アジアのロバのはるかに複雑な系統発生発達を指します。モンゴルのジグゲタイは、少なくとも3つの異なる被覆に分布しており、少なくとも2つのオナガーが分布しています。後者から、西線(コーカサス/イラン)は大部分が期限切れになっており、流通エリアの東部にある人はまだそれらを代表することができます。試験によると、ジゲゲタイの強い機能障害は、いくつかの移動波に関連付けられる可能性があります。一つの線はキアンの系統とより多く混合されているため、これはキアンが高い山の状態にもっと適応しているアジアのロバの形を表していることを示唆している可能性があります。 [30]
ステノニンの馬は、2.5〜300万年前にアジアで初めて証明されましたが、おそらく彼らはオープンするでしょう 馬のカミシ 北米に戻る。アジアのロバの密接な関係は不明ですが、更新世で形成される可能性があります Equus namadicus また 馬のsivalensis 見つけられた。 [21] 初期の化石 hemionus – ロシアのトロゴジのような動物は、100万年前後に伝えられています。 [最初に30] 化石亜種はほんの少し若いです E. H. nalaikhaensis モンゴルから、タウル川の堆積物から、約90万年前に磁気層序的に重要なジャラミロイベントに至るまで発見されました。 [32] と E. h。 binagadensis 中期および上部更新世から、別の比較的小さな唯一の化石のみが、アゼルバイジャンとイランで亜種を記録しました。 [4] タジキスタンからの発見は、チュジやオギ・キチクなど、若い更新世に伝えられています。特に中期および若い更新世では、アジアのロバはしばしばヨーロッパの野生のロバと一緒に個人的に発生しました( Hydruntinus ) の上。 [33] おそらく、更新世の終わりに絶滅したこのロバの形は、2017年の遺伝的研究によると、南および中央アジアの代表者とのつながりが強いため、アジアのロバの亜種としてのみ理解できます。 [30]
家畜化 [ 編集 | ソーステキストを編集します ]
現在のDNA検査では、今日の家の島がすべてアフリカのロバから降りてくることを確認しています。 DNAシーケンスに基づいて作成された家系図は、明らかにロバをアフリカとアジアの枝に分割します。後者には、アジアのロバが見つかります( 馬のhemionus )。アジアのロバが家畜化される可能性があるかどうか、そしてこれが過去に起こったかどうかの問題は議論の余地がありました。古いメソポタミア(URの標準)からの表現については、馬もロバもいない動物が認識されており、車の前でそれを伸ばすためにアジアのロバがシュメラーとアッカダーンによって飼いならされたことを少し急いでしました。 [34] しかし、最近の試みでは、彼らの恥ずかしさを人からアジアのロバに連れて行くことは決してできませんでした。一般に、アフリカのロバがメソポタミアでも飼いならされた可能性が高いと考えられています(先史時代の名前にもかかわらず、近東でも発生した)。
他の研究者は、シュメラーが家とアジアのロバの間の交差点も使用したと仮定しています。 [35] シリアのウムマラの青銅器時代の墓で見つかったコピーのゲノム(ADNA)の検査は、これがそのようなハイブリッドであることを示しています。関係する野生の形は、アジアのロバの地元の亜種であるシリアの半分でした。 [36]
脅威と保護 [ 編集 | ソーステキストを編集します ]
アジアのロバは、歴史的な時代にアジアのセミデザートを大いなる群れで交差させました。のどの渇きは、彼女を定期的に彼女の生息地のいくつかのウォーターポイントに導いた。マルコポーロは、13世紀に、アラビア、ペルシャ、トルケスタン、ゴビの動物の大規模な発生を報告しました。 [37] 今日の最大の脅威は、人間の居住地を広め、しばしば人を抱えていないが豊かな原材料の経済発展を広めることによる生息地の喪失です。アジアのロバは通常、牧草地や水源にアクセスするときに見ているため、大規模な農業との競争も大きな問題です。さらに、一方では、アジアのロバに対する違法な狩りは、皮のために、食物抽出の危険性が高まっています。 1990年代以降、在庫は52%減少しています。すべての亜種は、さまざまな程度ではありますが、脅かされています。全体の在庫はIUCN ASからのものです 脅迫された近く ( “潜在的に絶滅の危機にedeされています)。 [3]
からの保護対策に equidスペシャリストグループ IUCNは、アジアのロバの個々の個体群の第三国定住と第三国定住測定、生息地の使用、肉貿易の制御、地元住民の感作など、種類の生態学的ニーズと個々の亜種のさらなる調査を調整されています。 [3]
アナトールとシリアの半分 [ 編集 | ソーステキストを編集します ]

アナトリックのセミシンは、すでに古代で根絶されています。シリアの半室は、今日のイラクの地域で19世紀にまだ代表されていました。彼がますます一般的になった後、シリアの半分は第一次世界大戦でイギリス人とオスマン帝国の兵士にしばしば殺され、彼は絶滅の危機に陥りました。最後の野生のシリアの半分は1927年に殺されました。同じ年にウィーン動物園で1人の動物が死亡しました。
dschiggetai [ 編集 | ソーステキストを編集します ]
モンゴル人とも呼ばれるジゲゲイは、モンゴルの約41,900人の生きている人を持つ最も一般的な亜種であり、アジアのロバの総株の約4分の3です。そこの人口は安定した開発動向を示しています。モンゴル南部の大きなGobi-Aと大きなGobi-B保護エリアは、重要な保護地域です。 IUCNは、年間5〜10%の損失を想定しています。ジゲゲイは公式に保護されていますが、牧草地や水源の周りに遊牧的に生きている人口とその農業動物と多くの対立があります。密猟も1990年代から増加しており、これが株式の強い減少のもう1つの理由です。 [9] [6] 中国北部では、もう1つの約5000匹の動物が受け入れられており、カラミリ保護区で3,300人以上の動物が受け入れられています。もともとは、1980年代にここに住んでいた360のDschiggetaiしか住んでいませんでした。2000年からのみ、いくつかの大規模なグループがモンゴルから移住した後、その数が増加しました。 [3] [14] [38]
縮れ [ 編集 | ソーステキストを編集します ]

1960年代、Khurはイランとパキスタンの部分、およびその流通エリアのほとんどすべてのインド部分で死亡しました。彼は、インドのグジャラートにあるカッチェの小さな走りでしか生き残っていませんでした。ここでは、Dhrangadrhra-Wigressdatには珍しいロバの保護エリアがあります。 Khurは、アジアのロバの唯一の亜種であり、最近の過去に個人の数が着実に増加しています。 2014年には、4,000 Khureが小規模でカウントされました。それらの約30%は保護地域に住んでいます。それまでの間、動物はカッチチやブラックバック国立公園のビッグランなどの近隣地域にも広がります。人口は絶えず成長していますが、現時点では、ナルマダ運河は保護区の状態を危険にさらしています。なぜなら、淡水は多くの農民とその大型動物を引き付けているため、クーレ間の即時の競争であるからです。また、Kachchhの塩採掘の承認の要求もあります。 Chrusは、農業害虫や飼料競合他社の牛と見なされているため、保護地域の外の農家から撃たれます。 [3] [13]
セッション [ 編集 | ソーステキストを編集します ]

カザフスタンとトルクメニスタンの大部分でculanaが根絶された後、1940年代にソビエト連邦が50年間に5000人の動物の群れが形成されたBadkhys-Wildreservatを作成しました。トルクメニスタンのクラン在庫は最近縮小しました。1993年の5000人の動物のうち、2001年に推定580人です。トルクメニスタンの独立は密猟の世話をしました。残りの唯一の群れは、今日約420匹の動物が発生しているバッドキーズ自然保護区に一時的に住んでいました。トルクメニスタンで2番目に大きいグループは、おそらく350から400の動物があるサリカミシュセンケにあります。それまでの間、動物はまた、たとえばカプランキュルの自然保護区、バルサ・ケルムズ自然保護区、アルティン・エメル国立公園の他の場所で成功裏に頼りました。2014年の情報によると、後者は約2500から3000の動物を住んでいます。ただし、在庫は周辺地域の強力な農業使用によって制限されています。 IUCNは、Kulanの全体的な人口を最大2000人の成人と推定しています。この開発は、ソビエト連邦の時点で勝っていた効果的な狩猟管理のせいです。 [3] [39]
ヨーロッパ動物園協会(EAZA)のEEPの一部として、12人のヨーロッパ動物園が約50頭のコーランを維持しています。 EEPコーディネーターは、Krakow動物園のAnna Mekarskaです。 Kies Mine Landscape Weilbach(GRKW)の再栽培協会は、現在、自然環境でヨーロッパで6人の男性の会衆が半分のゲームを生きさせようとしています。
onager [ 編集 | ソーステキストを編集します ]

イラン北部の産業者の数は、約780人の個人に対して推定されています。 Onagerは2つの個別の集団に住んでいます。最大の人口は、約630人の動物を持つ14,000km²のトゥーラン保護エリアのうち導入され、2番目と小さい動物は3850km²で構成されるバーラムE-Goal保護エリアで発生し、ほぼ150人の個人があります。イラン政府は、密猟に対する厳しい罰と追加の水ポイントの作成を通じて、亜種の生存を確保しようとしています。 [3] [40]
欧州動物園協会(EAZA)の欧州保全育種プログラム(EEP)の一環として、約150人のオナージャーが今日科学的に主導の動物園に住んでいます。 EEPコーディネーターは、ハーゲンベック動物園のステファンヘリングハーゲンベックです。 ZoosのOnagerは、イスラエルのパワーラモン侵食クレーターのネゲブ砂漠にさらされましたが、人口の大部分は消費者との交差点から来ています。個体群のサイズは約250個の動物です。はるかに小さなグループがサウジアラビアにありました。 [3] [12番目]
パラスが使用する種の名前 hemionus ギリシャ語から派生しています ミミコン ヘミノス 何をオフ セミ DAME 「半分」と 熱意 ティニー 「ロバ」がまとめられます。指定 ヘミノス アナトリアとペルシャの動物のためにホーマーとアリストテレスの古代に使用されている場合、もともとミュールとラバと呼ばれていました。
名前の起源 onager 、ピーターボドダートが使用したものもギリシャ語です。 熱意 Tin -Tongue 、からの複合単語 熱意 ティニー 「ロバ」と 野生 ágrios “野生”。ラテン語の名前はそうです オナグラス また onager 。ギリシア人とローマ人はこれらの言葉をアフリカの野生のロバにのみ使用しましたが、シリアの半ベシドもvulgataで言及されています。
アチダリ 、 dschiggetai 、 gur-khar 、 縮れ と セッション アジアのロバの個々の個体群の局所的な一般名です。 [初め]
- Petra Kaczensky、Chris Walzer: アジアの野生のロバ – ゴビ砂漠の危険にさらされたサバイバルアーティスト。 の: コロン動物園のジャーナル。 51(3)、2008、S。147–163。
- 彼のクリンゲル: アフリカとアジアの野生のロバの社会組織と行動に関する観察(Equus africanus and Equus hemionus)。 の: Journal of Animal Psychology。 44、1975、S。323–331。
- P. Moehlman(hrsg。): equids:ゼブラ、ロバ、馬。ステータス調査と保全行動計画 。 IUCN、グランド、スイス2002。
- ロナルド・M・ノバック: ウォーカーの世界の哺乳類 。 Johns Hopkins University Press、1999、ISBN 0-8018-5789-9。
- ダニエルI.ルーベンシュタイン: 家族equidae(馬と親relative)。 In:Don E. Wilson、Russell A. Mittermeier(編): 世界の哺乳類のハンドブック。 第2巻: ひどい哺乳類。 Lynx Edicions、バルセロナ2011、ISBN 978-84-96553-77-4、S。104-143。
- ↑ a b c d そうです f g Colin Peter Groves、V。Mazák: アジアの野生のロバのいくつかの分類上の問題について。新しい亜種(ペリソダクタイラ、equidae)の説明を使用します。 の: 哺乳類の雑誌。 32、1967、S。321–355( [初め] )。
- ↑ a b Ingmar M Braun、Wolfgang Zessin: 旧石器時代の壁の芸術における馬の表現と、動物学的音像解釈を解釈しようとする試み。 の: ウルサス、動物園クラブと動物園シュヴェリンのニュースレター。 17(1)、2011年、S。4–26。
- ↑ a b c d そうです f g h 私 j k l P. Kaczensky、B。Lkhagvasuren、O。Pereladova、M。Hemami、A。Tilila: Equus hemionus。絶滅危species種のIUCNレッドリスト2020。 e.t7951a45171204( [2] ); 2020年12月10日に最後にアクセスしました。
- ↑ a b VéraIronmann、Marjan Mashhosour: Binagady(Azerbaidjan)とQazvin(Iran)の小さな等量:E。Hemionus binagadensis Nov。 subsp。およびE. Hydruntinus。 の: Geobios。 32(1)、1999、S。105–122。
- ↑ a b 彼のクリンゲル: アフリカとアジアの野生のロバの社会組織と行動に関する観察(Equus africanus and Equus hemionus)。 の: 応用動物行動科学。 60、1998、pp。103–113。 (再版、オリジナルの出版物 Journal of Animal Psychology。 44、1977、S。323–331)。
- ↑ a b c d そうです Petra Kaczensky、Chris Walzer: アジアの野生のロバ – ゴビ砂漠の危険にさらされたサバイバルアーティスト。 の: コロン動物園のジャーナル。 51(3)、2008、S。147–163。
- ↑ リチャード・P・リーディング、ヘンリー・M・ミックス、バダムジャビン・ラグヴァーヴァスレン、クラウディア・フェー、デビッド・P・ケイン、S。デュラムツセレン、スミヤイン・エンボルド: モンゴルのクーラン(Equus hemionus)のステータスと分布。 の: Zoological Societyのジャーナル。 254、2001、S。381–389。
- ↑ マーサI.グラインダー、ポールR.クラウスマン、ロバートS.ホフマン。 馬のロバ。 の: 哺乳類種。 794、2006、S。1–9。
- ↑ a b c d そうです f g h 私 C. Feh、B。Munkhtuya、S。Enkbold、T。Sukhbaatar: Gobi Khulan Equus Hemionus subspの生態と社会構造。モンゴルのゴビB国立公園で。 の: 生物学的保全。 101、2001、S。51–61。
- ↑ Petra Kaczensky、Ralph Kuehn、Badamjav Lhagvasures、Stephanie Pietsch、Weikang Yang、Chris Walzer: モンゴルのゴビにおけるアジアの野生のお尻の個体群の接続性。 の: 生物学的保全。 144、2011、S。920–929。
- ↑ G. Neumann-Denzau、H。Denzau: モンゴルの野生のお尻(Equus hemionus hemionus)の社会システムに関する発言。 の: モンゴルの生物資源に関する研究。 10、2007、S。177–187。
- ↑ a b デビッド・サルツ、メアリー・ローウェン、ダニエル・I・ルーベンシュタイン: 効果的な人口規模に対する再導入されたアジアの野生のロバの宇宙使用パターンの効果。 の: 保全生物学。 14(6)、2000、S。1852–1861。
- ↑ a b N.シャー、Q。Qureshi: Khur(Equus hemionus khur)の社会組織と空間分布。 の: モンゴルの生物資源に関する研究。 10、2007、S。189–200。
- ↑ a b Hu Defu、Zhang Daming und Lan Chunmei: 秋のカラマイラ除供給保護区におけるエクウスヘミオンとガゼラ亜尿素腫の生態習慣。 の: Journal of Forestry Research。 9(2)、1998、S。131–132。
- ↑ J.レンジャー、F。タタルチ、ch。 Walzer: アジアの野生のお尻のequus hemionusの摂食生態学。 の: モンゴルの生物資源に関する研究。 10、2007、S。93–97。
- ↑ S. P. Goyal、B。Sinha、N。Shah、H。S。Panwar: インドの野生のお尻に対するサルダル・サロバル・プロジェクトの脅威(Equus hemionus khur)。 の: 生物学的保全。 88、1999、S。277–284。
- ↑ デビッド・サルツ、ダニエル・I・ルーベンシュタイン: 再導入されたアジアの野生のお尻(Equus hemionus)の群れの個体群のダイナミクス。 の: 生態学的アプリケーション。 5(2)、1995、S。327–335。
- ↑ G. Neumann-Denzau: インドのリトルランオブカッチにおける野生のお尻(Equus hemionus khur)の誕生。 の: モンゴルの生物資源に関する研究。 10、2007、S。219–226。
- ↑ ヨハンナ・ペインター、ペトラ・カクゼンスキー、オユンサイカン・ガンバター、クラウス・フーバー、クリス・ウォルツァー: モンゴルの偉大なゴビ「B」厳密に保護された地域における同所性等量に関する比較寄生学的検査。 の: 欧州野生生物研究ジャーナル。 2010年 2:10.1007/s10344-010-0415-8 。
- ↑ ジュリア・T・ビルストラップ、アンダイン・セギン・オーランド、マティアス・スティラー、アウレリアン・ジノルハック、マアナサ・ラガヴァン、サンドラC.アピロ、ルドビッチオーランド: 現代および古代の等量のミトコンドリア系統発生体。 の: PLOS 1。 8(2)、2013年、S。E55950。
- ↑ a b そこにforstén: ミトコンドリア-DNAタイムテーブルとエクウスの進化:分子および古生物学的証拠の比較。 の: フェンニック動物学的アナレス。 28、1992、S。301–309。
- ↑ a b c サマンサ・A・プライス、オラフ・R・P・ビニンダ・エモンド: データの組み合わせを通じて、現存する馬、サイ、およびタピル(ペソダクタイラ)の包括的な系統発生。 の: Zoosystematics and Evolution。 85(2)、2009、S。277–292。
- ↑ Ariane Burke、Vera Eisenmann、Graeme K. Ambler: の体系的な位置 Hydruntinus 、更新世の絶滅種。 の: 第四紀研究。 59(3)、2003、S。459–469。
- ↑ Ludovic Orando、Dajansabsicour、Aranehe、Lrane、Christopher J. Custary、Véraine、Catherine Handsi: 絶滅の等量の地理的分布( Hydruntinus :哺乳類、equida)化石の形態学的および遺伝的分析によって明らかにされた。 の: 分子生態学。 15(8)、2006、S。2083–2093。
- ↑ a b ルドビッチ・オーランド、ジェシカ・L・メトカーフ、マリア・T・アルベルディ、ミゲル・テレス・アントゥネス、ドミニク・ボンジャン、マルセル・オッテ、ファビアナ・マーティン、ヴェラ・アイゼンマン、マージャン・マシュクール、フラビア・モレッロ、ジョセL.ウスティン、キャサリン・ヘンニ、アラン・クーパー: 古代のDNAを使用した等量の最近の進化の歴史を改訂します。 の: pnas。 106、2009、S。21754–21759。
- ↑ HákonJónsson、Mikkel Schubert、Andaine Seguin-Orlando、AurélienGinolhac、Lillian Petersen、Matteo Fumagallic、Anders Albrechtsen、Bent Petersen、Thorfinn S. Korneliussen、Julia T. Vilstrup、Teri Lear、Judsterup、Judsterup Med H. Alfarhan、Saleh A. Alquraishi、Khaled A. S. Al-Rasheid、Julia Stagegaard、GünterStrauss、Mads Frost Bertelsen、Thomas Sicheritz-Ponten、Douglas F. Antczak、Ernest Bailey、Rasmus Nielsen 広範な染色体可塑性にもかかわらず、等量での遺伝子流量の種分化。 の: pnas。 111(52)、2014、S。18655–18660。
- ↑ ピーター・サイモン・パラス: モンゴルのhemionusは、説明していると言った。 の: 新しい解説科学アカデミー帝国のペトロポリタナエ。 19、1774、S。394–414( [3] )。
- ↑ ダニエル・ルーベンシュタイン: 家族equidae(馬と親relative)。 In:Don E. Wilson、Russell A. Mittermeier(編): 世界の哺乳類のハンドブック。第2巻:ひづめの哺乳類。 Lynx Edicions、バルセロナ2011、ISBN 978-84-96553-77-4、S。104-143。
- ↑ Colin Groves、Peter Grubb: Ungulate Taxonomy。 ジョンズ・ホプキンス大学出版局、2011年、S。1–317(S。13–17)
- ↑ a b c E.アンドリュー・ベネット、ソフィー・シャンプロット、ジョリス・ピーターズ、ベンジャミン・S・アーバックル、シルビア・ギマレーズ、メラニー・プルヴォスト、シャーリ・バー・デイヴィッド、サイモン・J・M・デイビス、マシュー・ゴーティエ、ペトラ・カクゼンスキー、ラルフ・キューン、マージャン・マシュクル、アートゥル・モラル・マシュ・マシュ・マシュ・マシュ・マシュ・マシュ・マシュマン・エアプル・ア・マシュ・マシュ・マシュ・マシュ・マシュクル、 U、MietjeGermonpré、Can Y.Gündem、Mahmoud-èzaHemami、Pierre-ElieMoulé、AliyeÖtzan、Margarete Uerpmann、Chris Walzer、Thierry Grange、Eva-Maria Geigl: 古代および現代のDNAを通して、ユーラシアの野生のお尻の後期第四紀系統地理学を飼いならします。 の: PLOS 1。 12(4)、2017、S。E0174216、 doi:10.1371/journal.pone.0174216 。
- ↑ VéraIcernman: 鮮新世と更新世の等量:古生物学と分子生物学。 の: Senckenberg Research Instituteの宅配便業者。 256、2006、S。71–89。
- ↑ T. V. Kuznetsova、V。I。Zhegallo: 中央アジアの最も初期のクランであるモンゴルの更新世からのエクウス(hemionus)nalaikhaensis。 の: 古生物学ジャーナル。 43、2009、S。574–583。
- ↑ アン・フォルステン、シャリフ・シャラポフ: タジキスタンの新第三紀と更新世からの化石等(哺乳類、equidae)。 の: Geodiversitas。 22(2)、2000、S。293–314。
- ↑ P. ucko、G。dimbleby: 植物や動物の家畜化と搾取 。 Aldine Pub。、2007、ISBN 978-0-202-36169-7 (books.google.de) 。
- ↑ ジュリエット・クラットン・ブロック: 馬の力:人間社会における馬とロバの歴史。 1992、ISBN 0-674-406466-X。
- ↑ Eic Andrew Bennst、Jill Webサイト、Wedher-Benhofs、Gingrs Tei Range、Verry of Peter、Esloc: 初期の人間が作ったハイブリッド動物、シロメソポタミアのカンガスの遺伝的アイデンティティ。 の: 科学の進歩。 8、2022、S。EABM0218、 doi:10.1126/sciadv.abm0218 。
- ↑ Rohini Rangarajan: グジャラート州カッチの野生のお尻聖域における、インドの野生のお尻(Equus hemionus khur)の挙動に対する腸管寄生虫の影響。 マニパル大学、2008年。
- ↑ ヤン・ワイカン: 中国全体のエクウス・ヘミオスの状態に関する概要。 の: モンゴルの生物資源に関する研究。 10、2007、S。155–158。
- ↑ V:S。Lukarevskiy、Yu。 K. Goerlov: トルクメニスタンのクラン(エクウス・ヘミオス・パラス1775)。 の: モンゴルの生物資源に関する研究。 10、2007、S。231–240。
- ↑ L.タチン、B。F イランからの非常に危険にさらされているオナガー、エクウス・ヘミオナス・オナガーの行動と状態について今後。 の: モンゴルの生物資源に関する研究。 10、2007、S。253–259。
Recent Comments