
Trong sinh học tế bào, bộ máy trục chính (hoặc trục chính phân bào ) đề cập đến cấu trúc tế bào của các tế bào nhân chuẩn hình thành trong quá trình phân chia tế bào chị em giữa các tế bào con. Nó được gọi là trục chính phân bào trong quá trình nguyên phân, một quá trình tạo ra các tế bào con giống hệt nhau về mặt di truyền hoặc trục chính meo trong quá trình phân bào, một quá trình tạo ra giao tử với một nửa số lượng nhiễm sắc thể của tế bào cha.
Bên cạnh nhiễm sắc thể, bộ máy trục chính bao gồm hàng trăm protein. [1][2] Các vi ống bao gồm các thành phần phong phú nhất của máy móc.
Cấu trúc trục chính [ chỉnh sửa ]

Việc gắn các vi ống vào nhiễm sắc thể được trung gian bởi kinetochores, chủ động theo dõi sự hình thành trục chính và ngăn ngừa khởi phát phản vệ sớm. Microtubule trùng hợp và depolyme hóa ổ đĩa nhiễm sắc thể động lực. Sự phân hủy của các vi ống tạo ra sức căng tại kinetochores; [3] sự gắn kết lưỡng cực của kinetochores với các vi ống phát ra từ các cực của các tế bào đối diện với các lực căng thẳng, sắp xếp các nhiễm sắc thể ở xích đạo tế bào và phân chia chúng cho các tế bào con. Một khi mọi nhiễm sắc thể được định hướng hai chiều, anaphase bắt đầu và cohesin, mà cặp đôi nhiễm sắc thể chị em, bị cắt đứt, cho phép sự di chuyển của các nhiễm sắc thể chị em đến các cực đối diện.
Bộ máy trục chính của tế bào bao gồm các vi ống trục chính, các protein liên quan, bao gồm các động cơ phân tử kinesin và dynein, nhiễm sắc thể ngưng tụ và bất kỳ trung tâm hoặc aster nào có thể có ở các cực của trục chính tùy thuộc vào loại tế bào. [4] Thiết bị có hình elip mơ hồ trong mặt cắt ngang và thon ở phần cuối. Ở phần giữa rộng, được gọi là midzone trục chính, các vi ống đối cực được bó bởi kinesin. Ở đầu nhọn, được gọi là cực của trục chính, các vi ống được nhân lên bởi các trung tâm trong hầu hết các tế bào động vật. Acentrosomal hoặc anastral spindles thiếu centrosome hoặc asters ở các cực của trục chính, và xảy ra, ví dụ như trong bệnh teo cơ ở hầu hết các loài động vật. [5] bộ điều chỉnh chính của tổ chức và lắp ráp vi ống trục chính. Ở nấm, trục chính hình thành giữa các thân cực của trục chính được nhúng trong lớp vỏ hạt nhân, không bị phá vỡ trong quá trình nguyên phân.
Các protein liên quan đến vi ống và động lực học trục chính [ chỉnh sửa ]
Việc kéo dài và rút ngắn động của các vi ống trục chính, thông qua một quá trình được gọi là không ổn định động trục chính phân bào và thúc đẩy sự liên kết chính xác của nhiễm sắc thể ở midzone trục chính. Các protein liên kết với protein (MAP) liên kết với các vi ống ở midzone và các cực của trục chính để điều chỉnh động lực học của chúng. γ-tubulin là một biến thể tubulin chuyên biệt, tập hợp thành một phức hợp vòng gọi là-TuRC, tạo mầm cho quá trình trùng hợp của các heterodimers α / tubulin thành các vi ống. Việc tuyển dụng γ-TuRC vào khu vực màng ngoài tim giúp ổn định các đầu vi ống và neo chúng ở gần trung tâm tổ chức microtubule. Protein liên kết với vi ống mà Augmin hoạt động kết hợp với-TURC để tạo ra các vi ống mới. [6]
Phần cuối của vi ống đang phát triển được bảo vệ chống lại thảm họa bằng hoạt động của các protein theo dõi vi ống cộng đầu (+ TIPs) để thúc đẩy sự liên kết của chúng với kinetochores ở midzone. CLIP170 đã được hiển thị để bản địa hóa gần các đầu cộng microtubule trong các tế bào HeLa [7] và tích lũy trong kinetochores trong prometaphase. [8] Mặc dù CLIP170 nhận ra các đầu cộng của nó vẫn chưa rõ ràng, , [9][10] đề xuất vai trò của CLIP170 trong việc ổn định các đầu cộng và có thể làm trung gian gắn kết trực tiếp của chúng với kinetochores. [11] Các protein liên quan đến CLIP như CLASP1 ở người cũng đã được hiển thị để định vị các đầu cộng và cả kinetochore bên ngoài. như để điều chỉnh động lực học của vi ống kinetochore (Maiato 2003). Tương đồng CLASP trong Drosophila Xenopus và men được yêu cầu để lắp ráp trục chính thích hợp; ở động vật có vú, CLASP1 và CLASP2 đều góp phần vào sự lắp ráp trục chính và động lực học vi ống trong anaphase. [12] Sự trùng hợp đầu cuối có thể được kiểm duyệt thêm bởi protein EB1, liên kết trực tiếp các đầu phát triển của vi ống . [13] [14]
Đối lập với hoạt động của các protein ổn định microtubule này là một số yếu tố làm mất hoạt tính của microtubule. nhiễm sắc thể và đạt được lưỡng cực. Các siêu họ của kinesin-13 của MAP chứa một loại protein vận động có hướng kết thúc cộng với hoạt động khử phân hủy vi ống có liên quan bao gồm MCAK động vật có vú được nghiên cứu kỹ và Xenopus XKCM1. MCAK tập trung vào các mẹo phát triển của vi ống tại kinetochores, nơi nó có thể gây ra thảm họa khi cạnh tranh trực tiếp với hoạt động ổn định + TIP. [15] Những protein này khai thác năng lượng của quá trình thủy phân ATP để gây ra sự thay đổi cấu trúc của cấu trúc protofilament. [16] Mất hoạt động của chúng dẫn đến nhiều khiếm khuyết về phân bào. [15] Các protein gây mất ổn định vi ống bổ sung bao gồm Op18 / stathmin và katanin có vai trò trong việc tái cấu trúc trục chính phân bào cũng như thúc đẩy sự phân ly nhiễm sắc thể trong quá trình phản vệ.
Các hoạt động của các MAP này được điều chỉnh cẩn thận để duy trì động lực học vi ống thích hợp trong quá trình lắp ráp trục chính, với nhiều protein trong số đó đóng vai trò là chất nền kinase giống như Aurora và Polo. [17][18]
Tổ chức bộ máy trục chính [ chỉnh sửa ]
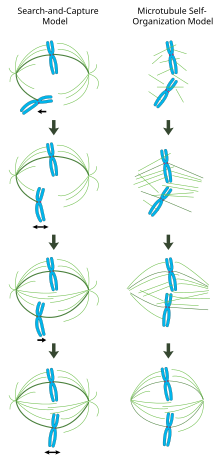
Trong một trục chính phân bào được hình thành chính xác, các nhiễm sắc thể định hướng hai chiều dọc theo đường xích đạo của tế bào với các vi ống trục chính được định hướng gần như vuông góc với nhiễm sắc thể, các đầu cộng của chúng được nhúng vào kinetochores và các đầu trừ của chúng được neo ở hai cực của tế bào. Sự định hướng chính xác của phức hợp này là cần thiết để đảm bảo sự phân chia nhiễm sắc thể chính xác và chỉ định mặt phẳng phân chia tế bào. Tuy nhiên, vẫn chưa rõ làm thế nào trục chính trở nên có tổ chức. Hai mô hình chiếm ưu thế trong lĩnh vực này, đó là hiệp đồng và không loại trừ lẫn nhau. Trong mô hình tìm kiếm và bắt giữ trục chính được tổ chức chủ yếu bằng cách phân tách các trung tâm tổ chức microtubule trung tâm (MTOC). Các vi ống trục chính phát ra từ các trung tâm và 'tìm kiếm' ra kinetochores; khi chúng liên kết một kinetochore, chúng trở nên ổn định và gây căng thẳng trên nhiễm sắc thể. Trong một mô hình thay thế tự lắp ráp các vi ống đã trải qua quá trình tạo mầm acentrosomal giữa các nhiễm sắc thể ngưng tụ. Bị ràng buộc bởi kích thước tế bào, các liên kết bên với các vi ống chống song song thông qua các protein vận động và các phần đính kèm cuối với kinetochores, các vi ống tự nhiên có cấu trúc giống như trục chính với các nhiễm sắc thể được xếp dọc theo đường xích đạo của tế bào.
Mô hình "tìm kiếm và bắt giữ" qua trung gian [ chỉnh sửa ]
Trong mô hình này, các vi ống được nhân lên tại các trung tâm tổ chức microtubule và trải qua sự phát triển nhanh chóng và thảm họa. tế bào chất cho kinetochores. Một khi chúng liên kết một kinetochore, chúng được ổn định và động lực của chúng bị giảm. Nhiễm sắc thể mới định hướng đơn sắc dao động trong không gian gần cực mà nó được gắn cho đến khi một vi ống từ cực đối diện liên kết với kinetochore chị em. Tập tin đính kèm thứ hai này tiếp tục ổn định tập tin đính kèm kinetochore vào trục chính phân bào. Dần dần, nhiễm sắc thể định hướng hai chiều được kéo về phía trung tâm của tế bào cho đến khi độ căng của vi ống được cân bằng ở cả hai phía của tâm động; nhiễm sắc thể sau đó dao động ở tấm metaphase cho đến khi khởi phát phản vệ giải phóng sự gắn kết của các nhiễm sắc thể chị em.
Trong mô hình này, các trung tâm tổ chức microtubule được định vị vào các cực của tế bào, sự phân tách của chúng được điều khiển bởi quá trình trùng hợp microtubule và 'trượt' của các vi ống trục chính chống song song với nhau ở giữa trục chính được trung gian bởi các lưỡng cực. [19][20] Các lực trượt như vậy có thể không chỉ giải thích cho việc tách cực của trục chính trong quá trình nguyên phân, mà còn kéo dài trục chính trong quá trình phản vệ muộn.
Tự tổ chức qua trung gian Chromatin của trục chính phân bào [ chỉnh sửa ]
Trái ngược với cơ chế tìm kiếm và bắt giữ trong đó các trung tâm chủ yếu chỉ đạo tổ chức của trục chính phân bào, mô hình này đề xuất rằng các vi ống được nhân lên một cách tự nhiên gần nhiễm sắc thể và tự lắp ráp thành các bó chống song song và áp dụng cấu trúc giống như trục chính. [21] Các thí nghiệm cổ điển của Heald và Karsenti cho thấy các trục và chức năng phân bào của hạt nhân xung quanh. Xenopus chiết xuất trứng và các mảng vi ống lưỡng cực được hình thành trong trường hợp không có centrosome và kinetochores. [22] Thật vậy, người ta cũng chứng minh rằng sự cắt bỏ tia laser của các centrosome trong các tế bào động vật có xương sống . [23] Theo sơ đồ này, hình dạng và kích thước của trục chính phân bào là một chức năng của các tính chất sinh lý của bảo vệ động cơ liên kết ngang eins. [24]
Tạo mầm vi ống qua trung gian Chromatin bằng gradient Ran GTP [ chỉnh sửa ]
Hệ số trao đổi nucleotide guanine cho GTPase Ran nhỏ (Điều hòa của nhiễm sắc thể 1) được gắn vào các nhiễm sắc thể thông qua các histone lõi H2A và H2B. [25] Do đó, một dải Ran liên kết với GTP được tạo ra xung quanh vùng lân cận của nhiễm sắc thể. Các hạt thủy tinh được phủ RCC1 tạo ra sự tạo mầm của vi ống và sự hình thành trục chính lưỡng cực trong Xenopus chiết xuất trứng, cho thấy chỉ riêng độ dốc Ran GTP là đủ để lắp ráp trục chính. [26] từ các tương tác ức chế thông qua các protein vận chuyển nhập khẩu β / α. Các SAF không liên kết sau đó thúc đẩy quá trình tạo mầm microtubule và ổn định xung quanh nhiễm sắc thể phân bào, và lưỡng cực trục chính được tổ chức bởi các protein động cơ microtubule. [27]
Điều chỉnh lắp ráp trục chính [ chỉnh sửa bởi các sự kiện phosphoryl hóa được xúc tác bởi kinase phân bào. Các phức kinase phụ thuộc cyclin (CDK) được kích hoạt bởi các cyclic phân bào, có sự dịch mã tăng lên trong quá trình nguyên phân. CDK1 (còn được gọi là CDC2) được coi là kinase phân bào chính trong các tế bào động vật có vú và được kích hoạt bởi Cyclin B1. Aurora kinase là cần thiết để lắp ráp và phân tách trục chính thích hợp. [28] Aurora Một liên kết với các trung tâm và được cho là để điều chỉnh sự xâm nhập của phân bào. Aurora B là một thành viên của tổ hợp hành khách nhiễm sắc thể và làm trung gian gắn kết nhiễm sắc thể-vi ống và sự gắn kết nhiễm sắc thể chị em. Kinase giống như polo, còn được gọi là PLK, đặc biệt là PLK1 có vai trò quan trọng trong việc duy trì trục chính bằng cách điều chỉnh động lực học vi ống. [29]
Cấu trúc nhiễm sắc thể phân bào [ chỉnh sửa ]
sự sao chép, các nhiễm sắc thể chị em được liên kết với nhau trong một khối vô định của DNA và protein rối rắm mà hầu như không thể phân chia thành từng tế bào con. Để tránh vấn đề này, mục nhập phân bào kích hoạt sự tái tổ chức mạnh mẽ của bộ gen được nhân đôi. Nhiễm sắc tố chị em được giải quyết và giải quyết lẫn nhau. Nhiễm sắc thể cũng rút ngắn chiều dài, lên tới 10.000 lần trong các tế bào động vật, [30] trong một quá trình gọi là ngưng tụ. Ngưng tụ bắt đầu trong tiên tri và nhiễm sắc thể được nén tối đa vào các cấu trúc hình que theo thời gian chúng được sắp xếp ở giữa trục chính tại metaphase. Điều này mang lại cho nhiễm sắc thể phân bào hình dạng cổ điển của X X thấy trong karyotypes, với mỗi nhiễm sắc thể chị em cô đặc được liên kết dọc theo chiều dài của chúng bằng protein cohesin và được nối, thường ở gần trung tâm, tại tâm động. [30] [31] [32]
Mặc dù các sắp xếp lại động này rất quan trọng để đảm bảo sự phân chia chính xác và độ chính xác cao của bộ gen, nhưng sự hiểu biết về cấu trúc nhiễm sắc thể của chúng vẫn chưa hoàn chỉnh. Tuy nhiên, một vài người chơi phân tử cụ thể đã được xác định: Topoisomerase II sử dụng quá trình thủy phân ATP để xúc tác quá trình khử các vướng mắc DNA, thúc đẩy phân giải chromatid chị em. [33] Condensin là phức hợp 5 tiểu đơn vị cũng sử dụng quá trình thủy phân ATP. Các thí nghiệm trong Xenopus Các chất chiết xuất từ trứng cũng liên quan đến chất liên kết Histone H1 như một bộ điều chỉnh quan trọng của quá trình nén nhiễm sắc thể phân bào. [35]
Điểm kiểm tra lắp ráp trục chính phân bào [ chỉnh sửa hoàn thành sự hình thành trục chính là một điểm chuyển tiếp quan trọng trong chu trình tế bào được gọi là điểm kiểm tra lắp ráp trục chính. Nếu nhiễm sắc thể không được gắn chính xác vào trục chính phân bào vào thời điểm của điểm kiểm tra này, sự khởi phát của phản vệ sẽ bị trì hoãn. [36] Thất bại của điểm kiểm tra lắp ráp trục chính này có thể dẫn đến vô hiệu hóa và có thể liên quan đến lão hóa và hình thành ung thư. [37]
Định hướng bộ máy trục chính [ chỉnh sửa ]
 Phim hoạt hình về tế bào biểu mô phân chia được bao quanh bởi mô biểu mô. Thiết bị trục chính quay bên trong tế bào. Sự quay vòng là kết quả của các vi ống thiên văn kéo theo các mối nối ba tế bào (TCJ), các trung tâm tín hiệu được định vị tại các khu vực nơi ba tế bào gặp nhau.
Phim hoạt hình về tế bào biểu mô phân chia được bao quanh bởi mô biểu mô. Thiết bị trục chính quay bên trong tế bào. Sự quay vòng là kết quả của các vi ống thiên văn kéo theo các mối nối ba tế bào (TCJ), các trung tâm tín hiệu được định vị tại các khu vực nơi ba tế bào gặp nhau.
Định hướng phân chia tế bào có tầm quan trọng lớn đối với kiến trúc mô, số phận tế bào và hình thái học. Các tế bào có xu hướng phân chia dọc theo trục dài của chúng theo quy tắc được gọi là quy tắc Hertwig. Trục phân chia tế bào được xác định bởi sự định hướng của bộ máy trục chính. Các tế bào phân chia dọc theo đường nối hai centrosome của bộ máy trục chính. Sau khi hình thành, bộ máy trục chính trải qua quá trình quay bên trong tế bào. Các vi ống thiên văn có nguồn gốc từ các trung tâm tiếp cận màng tế bào nơi chúng được kéo về phía các manh mối vỏ não cụ thể. In vitro sự phân bố các đầu mối vỏ được thiết lập bởi mô hình kết dính. [38] In vivo các dấu hiệu phân cực được xác định bằng cách định vị các mối nối Tricellular được định vị tại các đỉnh của tế bào. [39] các đầu mối vỏ não dẫn đến trường lực xác định hướng thiết bị trục chính cuối cùng và định hướng tiếp theo của sự phân chia tế bào.
Xem thêm [ chỉnh sửa ]
Tài liệu tham khảo [ chỉnh sửa ]
- ^ C. E. Walczak; R. Trời (2008). "Các cơ chế của hội và trục chính phân bào". Tạp chí quốc tế về tế bào học . 265 : 111 Ảo158. doi: 10.1016 / s0074-7696 (07) 65003-7.
- ^ Helmke KJ, Heald R, Wilbur JD (2013). "Tương tác giữa kiến trúc trục chính và chức năng". Mục sư Mol. Biol . 306 : 83 Ảo125. doi: 10.1016 / B978-0-12-407694-5.00003-1. PMID 24016524.
- ^ E. Vô danh; V. H. Ramey (ngày 1 tháng 11 năm 2009). "Cấu trúc chức năng hiểu biết sâu sắc về phức hợp men Dam1 kinetochore". J Cell Sci . 122 : 3831 Từ3836. doi: 10.1242 / jcs.004689. PMC 2773187 .
- ^ Campbell, Neil A.; Jane B. Reece (2005). Sinh học, tái bản lần thứ 7 . San Francisco: Benjamin Cummings. tr.222222222. Sđt 0-8053-7171-0.
- ^ Manandhar Gf; Schatten H; Sutovsky P (2005). "Giảm trung tâm trong quá trình phát sinh giao tử và ý nghĩa của nó". Biol. Reprod . 72 (1): 2 trận13. doi: 10.1095 / biolreprod.104.031245. PMID 15385423.
- ^ Petry S, et al. (2013). "Tạo mầm vi ống phân nhánh trong chiết xuất trứng Xenopus qua trung gian bởi augmin và TPX2". Tế bào . 152 : 768 Mạnh777. doi: 10.1016 / j.cell.2012.12.044. PMC 3680448 . PMID 23415226.
- ^ J.E. Bệnh sốt rét; T.E. Kreis (1990). "Xác định một protein liên kết vi ống nhạy cảm với nucleotide mới trong các tế bào HeLa". Bi Cell J . 110 (5): 1623 Tiết1633. doi: 10.1083 / jcb.110.5.1623. PMC 2200191 . PMID 1970824.
- ^ D. Dujardin; Hoa Kỳ Wacker; A. Moreau; T.A. Schroer; J.E Rickard; J.R. DeMey (1998). "Bằng chứng cho vai trò của CLIP-170 trong việc thiết lập sự liên kết nhiễm sắc thể của metaphase". Bi Cell J . 141 (4): 849 Từ862. doi: 10.1083 / jcb.141.4.849. PMC 2132766 . PMID 9585405.
- ^ D. Ngăm ngăm; P. Y tá (2000). "CLIP-170 giống như tip1p tổ chức không gian động lực học vi ống trong nấm men phân hạch". Tế bào . 102 (5): 695 Điêu704. doi: 10.1016 / S0092-8674 (00) 00091-X. PMID 11007487.
- ^ Y.A. Komarova; NHƯ. Kojima; et al. (2002). "Các protein liên kết tế bào chất thúc đẩy giải cứu vi ống trong cơ thể". Bi Cell J . 159 : 589 Từ599. doi: 10.1083 / jcb.200208058. PMC 2173097 . PMID 12446741.
- ^ S. Đá vàng; C. Reyes; G. Gay; T. Courthéoux; M. Dubarry; et al. (2010). "Protein Tip1 / CLIP-170 là cần thiết cho sự di chuyển chính xác của nhiễm sắc thể trong nấm men phân hạch". PLoS ONE . 5 : e10634. doi: 10.1371 / tạp chí.pone.0010634. PMC 2869355 . PMID 20498706.
- ^ A.L. Pereira; A.J. Pereira; A.R.R. Maia; et al. (1 tháng 10 năm 2006). "Động vật có vú CLASP1 và CLASP2 hợp tác để đảm bảo tính trung thực của phân bào bằng cách điều chỉnh chức năng trục chính và Kinetochore". Tế bào Mol Biol . 17 : 4526 Từ4542. doi: 10.1091 / mbc.E06-07-0579. PMC 1635371 . PMID 16914514.
- ^ A. Akhmanova; M.O. Steinmetz (tháng 4 năm 2008). "Theo dõi kết thúc: một mạng lưới protein động kiểm soát số phận của các mẹo vi ống". Nat Rev Mol Cell Biol . 9 (4): 309 Từ 322. doi: 10.1038 / nrm2369. PMID 18322465.
- ^ J.S. Tirnauer; S. Grego; E.D. Cá hồi; T.J. Mitchison (1 tháng 10 năm 2002). "Tương tác EB1-microtubule trong chiết xuất trứng Xenopus: Vai trò của EB1 trong ổn định vi ống và cơ chế nhắm mục tiêu đến vi ống". Tế bào Mol Biol . 13 (10): 3614 Tiết3626. doi: 10.1091 / mbc.02-04-0210. PMC 129970 . PMID 12388761.
- ^ a b M.E. Tanenbaum; R.H. Medema; A. Akhmanova (2011). "Quy định nội địa hóa và hoạt động của microtubule depolymerase MCAK". Kiến trúc sinh học . 1 (2): 80 Ném87. doi: 10,4161 / sinh học.1.2.15807. PMC 3158623 . PMID 21866268.
- ^ H. Niederstrasser; H. Salehi-Had; E.C. Gan; C. Walczak; E. Nogales (2002). "XKCM1 hoạt động trên một protofilament duy nhất và yêu cầu đầu cuối C của tubulin". J Mol Biol . 316 (3): 817 Ảo828. doi: 10.1006 / jmbi.2001.5360. PMID 11866534.
- ^ a b H. Cà chua; P Sampaio; C.Em Sunkel (2004). "Các protein liên quan đến vi ống và vai trò thiết yếu của chúng trong quá trình nguyên phân". Int Rev Cytol . 241 : 53 trừ153. doi: 10.1016 / S0074-7696 (04) 41002-X. PMID 15548419.
- ^ R. Tournebize; A. Popov; K. Kinoshita; A.J. Ashford; et al. (2000). "Kiểm soát động lực học vi ống bằng các hoạt động đối kháng của XMAP215 và XKCM1 trong chiết xuất trứng Xenopus". Nat Cell Biol . 2 : 13 Ảo19. đổi: 10.1038 / 71330. PMID 10620801.
- ^ J. McIntosh; S.C. Landis (1971). "Sự phân bố của các vi ống trục chính trong quá trình nguyên phân trong tế bào người được nuôi cấy". Bi Cell J . 49 (2): 468 Tiết497. doi: 10.1083 / jcb.49.2.468. PMC 2108320 . PMID 19866774.
- ^ D.J. Nhọn; K.L. McDonald; H.M. Nâu; et al. (1999). "Các kinesin lưỡng cực, CLP61F, các vi ống liên kết chéo trong các bó vi ống giữa các cực của phôi bào phân bào phôi Drosophila". Bi Cell J . 144 (1): 125 Ảo138. doi: 10.1083 / jcb.144.1.125. PMC 2148119 . PMID 9885249.
- ^ M.A. Hallen; S.A. Endow (2009). "Lắp ráp trục chính Anastral: một mô hình toán học". Biophys J . 97 (8): 2191 Tiết2201. doi: 10.1016 / j.bpj.2009.08.008. PMC 2764103 . PMID 19843451.
- ^ R. Trời cao; R. Tournebize; et al. (1996). "Tự tổ chức các vi ống thành các trục lưỡng cực xung quanh nhiễm sắc thể nhân tạo trong chiết xuất trứng Xenopus". Thiên nhiên . 382 (6590): 420 Chiếc425. doi: 10.1038 / 382420a0. PMID 8684481.
- ^ A. Khodjakov; R.W Cole; B.R. Sồi; C.L. Rieder (2000). "Sự hình thành trục chính phân bào độc lập Centrosome ở động vật có xương sống". Curr Biol . 10 (2): 59 Tái67. doi: 10.1016 / S0960-9822 (99) 00276-6. PMID 10662665.
- ^ K.S. Burbank; T.J. Mitchison; D.S Fisher (2007). "Các mô hình trượt và cụm cho lắp ráp trục chính". Curr Biol . 17 (16): 1373 Ảo1383. doi: 10.1016 / j.cub.2007.07.058. PMID 17702580.
- ^ Makde R, Anh J, Yennawar H, Tan S (2010). "Cấu trúc của yếu tố nhiễm sắc RCC1 liên kết với hạt nhân nucleosome". Thiên nhiên . 467 (7315): 562 Tắt566. doi: 10.1038 / thiên nhiên09321. PMC 3168546 . PMID 20739938.
- ^ Halpin D, Kalab P, Wang J, Weis K, Heald R (2011). "Lắp ráp trục chính phân bào xung quanh các hạt được phủ RCC1 trong chiết xuất trứng Xenopus". PLoS Biol . 9 (12): e1001225. doi: 10.1371 / tạp chí.pbio.1001225. PMC 3246454 . PMID 22215983.
- ^ Fu J, Jiang Q, Zhang C (2010). "Phối hợp các sự kiện chu trình tế bào của Ran GTPase". Giáo dục thiên nhiên . 3 (9): 32.
- ^ A.R. Thùng; F. Gergely (2007). "Aurora A: Nhà sản xuất và phá vỡ các trục chính". J Cell Sci . 120 : 2987 2122996. doi: 10.1242 / jcs.013136.
- ^ Peters, U., J. Cherian; et al. (2006). "Kiểm tra không gian kiểu hình phân chia tế bào và chức năng kinase giống như Polo sử dụng các phân tử nhỏ". Nat Chem Biol . 2 (11): 618 Điêu26. doi: 10.1038 / nchembio826. PMID 17028580. CS1 duy trì: Nhiều tên: danh sách tác giả (liên kết)
- ^ a b Morgan DO: The Cell Chu kỳ: Nguyên tắc của Kiểm soát (Primers inBiology) London: New Science Press Ltd; 2007: 297. ISBN 976-0-9539181-2-6
- ^ Belmont AS (2010). "Tổ chức chromatin quy mô lớn: Tốt, đáng ngạc nhiên và vẫn còn bối rối". Sinh học tế bào Curr Opin . 26 : 69 bù78. doi: 10.1016 / j.ceb.2013.10.002. PMC 3927141 . PMID 24529248.
- ^ Marko, JF. Nhiễm sắc thể phân bào: cấu trúc và cơ học. 2012. Tổ chức và chức năng bộ gen trong nhân tế bào. Wiley-VCH, Ch. 18, 449-485. doi: 10.1002 / 9783527639991.ch18
- ^ Champoux JJ (2001). "TOPOISOMERASES DNA: Cấu trúc, chức năng và cơ chế". Annu Rev Biochem . 70 (1): 369 Từ413. doi: 10.1146 / annurev.biochem70.1.369. PMID 11395412.
- ^ Hirano T (2012). "Condensins: tổ chức phổ biến của nhiễm sắc thể với các chức năng đa dạng". Gen Dev . 26 (15): 1659 Tiết1678. doi: 10.1101 / gad.194746.112. PMC 3418584 . PMID 22855829.
- ^ Maresca TJ, Freedman BS, Heald R (2005). "Histone H1 rất cần thiết cho kiến trúc nhiễm sắc thể phân bào và sự phân ly trong chiết xuất trứng Xenopus laevis". J. Biol tế bào . 169 (6): 859 Ảo69. doi: 10.1083 / jcb.200503031. PMC 2171634 . PMID 15967810.
- ^ Raven, Peter H.; Ray F. Evert; Susan E. Eichhorn (2005). Sinh học thực vật, tái bản lần thứ 7 . New York: W.H. Freeman và Nhà xuất bản Công ty. tr. 59. ISBN 0-7167-1007-2.
- ^ Baker DJ, Chen J, van Deursen JM (2005). "Điểm kiểm tra phân bào trong ung thư và lão hóa: chuột đã dạy chúng ta điều gì?". Curr. Ý kiến Biol tế bào . 17 (6): 583 Tắt9. doi: 10.1016 / j.ceb.2005.09.011. PMID 16226453.
- ^ Thery M, Jimenez-Dalmaroni A, Racine V, Bornens M, Jul Rich F (2007). "Nghiên cứu thực nghiệm và lý thuyết về định hướng trục chính phân bào". Thiên nhiên . 447 : 493 Tắt6. doi: 10.1038 / thiên nhiên05786. PMID 17495931.
- ^ Bosveld F, Markova O, Guirao B, Martin C, Wang Z, Pierre A, Balakireva M, Gaugue I, Ainslie A, Barshorou N, Lubensky DK, Minc N, Bellaïche ). "Các mối nối ba biểu mô hoạt động như các cảm biến hình dạng tế bào xen kẽ để định hướng nguyên phân". Thiên nhiên . 530 : 496 Ảo8. doi: 10.1038 / thiên nhiên16970. PMID 26886796.
Liên kết ngoài [ chỉnh sửa ]
Định hướng bộ máy trục chính [ chỉnh sửa ]
Định hướng phân chia tế bào có tầm quan trọng lớn đối với kiến trúc mô, số phận tế bào và hình thái học. Các tế bào có xu hướng phân chia dọc theo trục dài của chúng theo quy tắc được gọi là quy tắc Hertwig. Trục phân chia tế bào được xác định bởi sự định hướng của bộ máy trục chính. Các tế bào phân chia dọc theo đường nối hai centrosome của bộ máy trục chính. Sau khi hình thành, bộ máy trục chính trải qua quá trình quay bên trong tế bào. Các vi ống thiên văn có nguồn gốc từ các trung tâm tiếp cận màng tế bào nơi chúng được kéo về phía các manh mối vỏ não cụ thể. In vitro sự phân bố các đầu mối vỏ được thiết lập bởi mô hình kết dính. [38] In vivo các dấu hiệu phân cực được xác định bằng cách định vị các mối nối Tricellular được định vị tại các đỉnh của tế bào. [39] các đầu mối vỏ não dẫn đến trường lực xác định hướng thiết bị trục chính cuối cùng và định hướng tiếp theo của sự phân chia tế bào.
Xem thêm [ chỉnh sửa ]
Tài liệu tham khảo [ chỉnh sửa ]
- ^ C. E. Walczak; R. Trời (2008). "Các cơ chế của hội và trục chính phân bào". Tạp chí quốc tế về tế bào học . 265 : 111 Ảo158. doi: 10.1016 / s0074-7696 (07) 65003-7.
- ^ Helmke KJ, Heald R, Wilbur JD (2013). "Tương tác giữa kiến trúc trục chính và chức năng". Mục sư Mol. Biol . 306 : 83 Ảo125. doi: 10.1016 / B978-0-12-407694-5.00003-1. PMID 24016524.
- ^ E. Vô danh; V. H. Ramey (ngày 1 tháng 11 năm 2009). "Cấu trúc chức năng hiểu biết sâu sắc về phức hợp men Dam1 kinetochore". J Cell Sci . 122 : 3831 Từ3836. doi: 10.1242 / jcs.004689. PMC 2773187 .
- ^ Campbell, Neil A.; Jane B. Reece (2005). Sinh học, tái bản lần thứ 7 . San Francisco: Benjamin Cummings. tr.222222222. Sđt 0-8053-7171-0.
- ^ Manandhar Gf; Schatten H; Sutovsky P (2005). "Giảm trung tâm trong quá trình phát sinh giao tử và ý nghĩa của nó". Biol. Reprod . 72 (1): 2 trận13. doi: 10.1095 / biolreprod.104.031245. PMID 15385423.
- ^ Petry S, et al. (2013). "Tạo mầm vi ống phân nhánh trong chiết xuất trứng Xenopus qua trung gian bởi augmin và TPX2". Tế bào . 152 : 768 Mạnh777. doi: 10.1016 / j.cell.2012.12.044. PMC 3680448 . PMID 23415226.
- ^ J.E. Bệnh sốt rét; T.E. Kreis (1990). "Xác định một protein liên kết vi ống nhạy cảm với nucleotide mới trong các tế bào HeLa". Bi Cell J . 110 (5): 1623 Tiết1633. doi: 10.1083 / jcb.110.5.1623. PMC 2200191 . PMID 1970824.
- ^ D. Dujardin; Hoa Kỳ Wacker; A. Moreau; T.A. Schroer; J.E Rickard; J.R. DeMey (1998). "Bằng chứng cho vai trò của CLIP-170 trong việc thiết lập sự liên kết nhiễm sắc thể của metaphase". Bi Cell J . 141 (4): 849 Từ862. doi: 10.1083 / jcb.141.4.849. PMC 2132766 . PMID 9585405.
- ^ D. Ngăm ngăm; P. Y tá (2000). "CLIP-170 giống như tip1p tổ chức không gian động lực học vi ống trong nấm men phân hạch". Tế bào . 102 (5): 695 Điêu704. doi: 10.1016 / S0092-8674 (00) 00091-X. PMID 11007487.
- ^ Y.A. Komarova; NHƯ. Kojima; et al. (2002). "Các protein liên kết tế bào chất thúc đẩy giải cứu vi ống trong cơ thể". Bi Cell J . 159 : 589 Từ599. doi: 10.1083 / jcb.200208058. PMC 2173097 . PMID 12446741.
- ^ S. Đá vàng; C. Reyes; G. Gay; T. Courthéoux; M. Dubarry; et al. (2010). "Protein Tip1 / CLIP-170 là cần thiết cho sự di chuyển chính xác của nhiễm sắc thể trong nấm men phân hạch". PLoS ONE . 5 : e10634. doi: 10.1371 / tạp chí.pone.0010634. PMC 2869355 . PMID 20498706.
- ^ A.L. Pereira; A.J. Pereira; A.R.R. Maia; et al. (1 tháng 10 năm 2006). "Động vật có vú CLASP1 và CLASP2 hợp tác để đảm bảo tính trung thực của phân bào bằng cách điều chỉnh chức năng trục chính và Kinetochore". Tế bào Mol Biol . 17 : 4526 Từ4542. doi: 10.1091 / mbc.E06-07-0579. PMC 1635371 . PMID 16914514.
- ^ A. Akhmanova; M.O. Steinmetz (tháng 4 năm 2008). "Theo dõi kết thúc: một mạng lưới protein động kiểm soát số phận của các mẹo vi ống". Nat Rev Mol Cell Biol . 9 (4): 309 Từ 322. doi: 10.1038 / nrm2369. PMID 18322465.
- ^ J.S. Tirnauer; S. Grego; E.D. Cá hồi; T.J. Mitchison (1 tháng 10 năm 2002). "Tương tác EB1-microtubule trong chiết xuất trứng Xenopus: Vai trò của EB1 trong ổn định vi ống và cơ chế nhắm mục tiêu đến vi ống". Tế bào Mol Biol . 13 (10): 3614 Tiết3626. doi: 10.1091 / mbc.02-04-0210. PMC 129970 . PMID 12388761.
- ^ a b M.E. Tanenbaum; R.H. Medema; A. Akhmanova (2011). "Quy định nội địa hóa và hoạt động của microtubule depolymerase MCAK". Kiến trúc sinh học . 1 (2): 80 Ném87. doi: 10,4161 / sinh học.1.2.15807. PMC 3158623 . PMID 21866268.
- ^ H. Niederstrasser; H. Salehi-Had; E.C. Gan; C. Walczak; E. Nogales (2002). "XKCM1 hoạt động trên một protofilament duy nhất và yêu cầu đầu cuối C của tubulin". J Mol Biol . 316 (3): 817 Ảo828. doi: 10.1006 / jmbi.2001.5360. PMID 11866534.
- ^ a b H. Cà chua; P Sampaio; C.Em Sunkel (2004). "Các protein liên quan đến vi ống và vai trò thiết yếu của chúng trong quá trình nguyên phân". Int Rev Cytol . 241 : 53 trừ153. doi: 10.1016 / S0074-7696 (04) 41002-X. PMID 15548419.
- ^ R. Tournebize; A. Popov; K. Kinoshita; A.J. Ashford; et al. (2000). "Kiểm soát động lực học vi ống bằng các hoạt động đối kháng của XMAP215 và XKCM1 trong chiết xuất trứng Xenopus". Nat Cell Biol . 2 : 13 Ảo19. đổi: 10.1038 / 71330. PMID 10620801.
- ^ J. McIntosh; S.C. Landis (1971). "Sự phân bố của các vi ống trục chính trong quá trình nguyên phân trong tế bào người được nuôi cấy". Bi Cell J . 49 (2): 468 Tiết497. doi: 10.1083 / jcb.49.2.468. PMC 2108320 . PMID 19866774.
- ^ D.J. Nhọn; K.L. McDonald; H.M. Nâu; et al. (1999). "Các kinesin lưỡng cực, CLP61F, các vi ống liên kết chéo trong các bó vi ống giữa các cực của phôi bào phân bào phôi Drosophila". Bi Cell J . 144 (1): 125 Ảo138. doi: 10.1083 / jcb.144.1.125. PMC 2148119 . PMID 9885249.
- ^ M.A. Hallen; S.A. Endow (2009). "Lắp ráp trục chính Anastral: một mô hình toán học". Biophys J . 97 (8): 2191 Tiết2201. doi: 10.1016 / j.bpj.2009.08.008. PMC 2764103 . PMID 19843451.
- ^ R. Trời cao; R. Tournebize; et al. (1996). "Tự tổ chức các vi ống thành các trục lưỡng cực xung quanh nhiễm sắc thể nhân tạo trong chiết xuất trứng Xenopus". Thiên nhiên . 382 (6590): 420 Chiếc425. doi: 10.1038 / 382420a0. PMID 8684481.
- ^ A. Khodjakov; R.W Cole; B.R. Sồi; C.L. Rieder (2000). "Sự hình thành trục chính phân bào độc lập Centrosome ở động vật có xương sống". Curr Biol . 10 (2): 59 Tái67. doi: 10.1016 / S0960-9822 (99) 00276-6. PMID 10662665.
- ^ K.S. Burbank; T.J. Mitchison; D.S Fisher (2007). "Các mô hình trượt và cụm cho lắp ráp trục chính". Curr Biol . 17 (16): 1373 Ảo1383. doi: 10.1016 / j.cub.2007.07.058. PMID 17702580.
- ^ Makde R, Anh J, Yennawar H, Tan S (2010). "Cấu trúc của yếu tố nhiễm sắc RCC1 liên kết với hạt nhân nucleosome". Thiên nhiên . 467 (7315): 562 Tắt566. doi: 10.1038 / thiên nhiên09321. PMC 3168546 . PMID 20739938.
- ^ Halpin D, Kalab P, Wang J, Weis K, Heald R (2011). "Lắp ráp trục chính phân bào xung quanh các hạt được phủ RCC1 trong chiết xuất trứng Xenopus". PLoS Biol . 9 (12): e1001225. doi: 10.1371 / tạp chí.pbio.1001225. PMC 3246454 . PMID 22215983.
- ^ Fu J, Jiang Q, Zhang C (2010). "Phối hợp các sự kiện chu trình tế bào của Ran GTPase". Giáo dục thiên nhiên . 3 (9): 32.
- ^ A.R. Thùng; F. Gergely (2007). "Aurora A: Nhà sản xuất và phá vỡ các trục chính". J Cell Sci . 120 : 2987 2122996. doi: 10.1242 / jcs.013136.
- ^ Peters, U., J. Cherian; et al. (2006). "Kiểm tra không gian kiểu hình phân chia tế bào và chức năng kinase giống như Polo sử dụng các phân tử nhỏ". Nat Chem Biol . 2 (11): 618 Điêu26. doi: 10.1038 / nchembio826. PMID 17028580. CS1 duy trì: Nhiều tên: danh sách tác giả (liên kết)
- ^ a b Morgan DO: The Cell Chu kỳ: Nguyên tắc của Kiểm soát (Primers inBiology) London: New Science Press Ltd; 2007: 297. ISBN 976-0-9539181-2-6
- ^ Belmont AS (2010). "Tổ chức chromatin quy mô lớn: Tốt, đáng ngạc nhiên và vẫn còn bối rối". Sinh học tế bào Curr Opin . 26 : 69 bù78. doi: 10.1016 / j.ceb.2013.10.002. PMC 3927141 . PMID 24529248.
- ^ Marko, JF. Nhiễm sắc thể phân bào: cấu trúc và cơ học. 2012. Tổ chức và chức năng bộ gen trong nhân tế bào. Wiley-VCH, Ch. 18, 449-485. doi: 10.1002 / 9783527639991.ch18
- ^ Champoux JJ (2001). "TOPOISOMERASES DNA: Cấu trúc, chức năng và cơ chế". Annu Rev Biochem . 70 (1): 369 Từ413. doi: 10.1146 / annurev.biochem70.1.369. PMID 11395412.
- ^ Hirano T (2012). "Condensins: tổ chức phổ biến của nhiễm sắc thể với các chức năng đa dạng". Gen Dev . 26 (15): 1659 Tiết1678. doi: 10.1101 / gad.194746.112. PMC 3418584 . PMID 22855829.
- ^ Maresca TJ, Freedman BS, Heald R (2005). "Histone H1 rất cần thiết cho kiến trúc nhiễm sắc thể phân bào và sự phân ly trong chiết xuất trứng Xenopus laevis". J. Biol tế bào . 169 (6): 859 Ảo69. doi: 10.1083 / jcb.200503031. PMC 2171634 . PMID 15967810.
- ^ Raven, Peter H.; Ray F. Evert; Susan E. Eichhorn (2005). Sinh học thực vật, tái bản lần thứ 7 . New York: W.H. Freeman và Nhà xuất bản Công ty. tr. 59. ISBN 0-7167-1007-2.
- ^ Baker DJ, Chen J, van Deursen JM (2005). "Điểm kiểm tra phân bào trong ung thư và lão hóa: chuột đã dạy chúng ta điều gì?". Curr. Ý kiến Biol tế bào . 17 (6): 583 Tắt9. doi: 10.1016 / j.ceb.2005.09.011. PMID 16226453.
- ^ Thery M, Jimenez-Dalmaroni A, Racine V, Bornens M, Jul Rich F (2007). "Nghiên cứu thực nghiệm và lý thuyết về định hướng trục chính phân bào". Thiên nhiên . 447 : 493 Tắt6. doi: 10.1038 / thiên nhiên05786. PMID 17495931.
- ^ Bosveld F, Markova O, Guirao B, Martin C, Wang Z, Pierre A, Balakireva M, Gaugue I, Ainslie A, Barshorou N, Lubensky DK, Minc N, Bellaïche ). "Các mối nối ba biểu mô hoạt động như các cảm biến hình dạng tế bào xen kẽ để định hướng nguyên phân". Thiên nhiên . 530 : 496 Ảo8. doi: 10.1038 / thiên nhiên16970. PMID 26886796.
Liên kết ngoài [ chỉnh sửa ]