Mosasaurs (từ tiếng Latin Mosa có nghĩa là 'sông Meuse', và Hy Lạp 1945σύρύρύρ có nghĩa là 'thằn lằn') là một nhóm tuyệt chủng của các loài bò sát biển lớn có tổng cộng 38 chi. Hóa thạch đầu tiên của chúng được phát hiện trong một mỏ đá vôi tại Maastricht trên Meuse vào năm 1764. Mosasaur có lẽ đã tiến hóa từ một nhóm thằn lằn thủy sinh đã tuyệt chủng [1] được gọi là aigializard trong kỷ Phấn trắng sớm. Trong suốt 20 triệu năm cuối của kỷ Phấn trắng (thời kỳ Turian-Maastrichtian), với sự tuyệt chủng của loài ichthyizard và pliosaur, mosasaur trở thành loài săn mồi biển thống trị. Chúng bị tuyệt chủng do sự kiện K-PG vào cuối thời kỳ kỷ Phấn trắng, khoảng 66 triệu năm trước.
Mô tả [ chỉnh sửa ]


Mosasaur hít thở không khí, là những người bơi lội mạnh mẽ và thích nghi tốt với cuộc sống ở vùng biển nội địa nông, ấm áp phổ biến trong thời kỳ cuối kỷ Phấn trắng. Mosasaur thích nghi tốt với môi trường này đến nỗi chúng sinh ra để sống trẻ, thay vì quay trở lại bờ để đẻ trứng như rùa biển. [2]
Mosasaur được biết đến nhỏ nhất là Dallasaurus Turneri đó là dài dưới 1 m (3,3 ft). Loài mosasaur lớn hơn là điển hình hơn, với nhiều loài phát triển dài hơn 4 m (13 ft). Mosasaurus hoffmannii loài lớn nhất được biết đến, có thể đã đạt tới chiều dài 17 m (56 ft). [3] Hiện tại, bộ xương mosasaur được trưng bày công khai lớn nhất trên thế giới đang được trưng bày tại Khám phá Fossil Canada Trung tâm ở Morden, Manitoba. Mẫu vật có biệt danh "Bruce", chỉ dài hơn 13 m (43 ft). [4]
Mosasaur có hình dạng cơ thể tương tự như thằn lằn theo dõi thời hiện đại (varanids), nhưng dài hơn và được sắp xếp hợp lý để bơi lội. Xương chi của chúng bị giảm chiều dài và mái chèo của chúng được hình thành bằng cách nối giữa ngón tay dài và xương ngón chân. Đuôi của chúng rất rộng và cung cấp năng lượng đầu máy. Cho đến gần đây, mosasaur được cho là đã bơi trong một phương pháp tương tự như phương pháp được sử dụng ngày nay bởi cá chình và rắn biển, nhấp nhô toàn bộ cơ thể của chúng từ bên này sang bên kia. Tuy nhiên, bằng chứng mới cho thấy nhiều loài mosasaur tiên tiến có sán lớn hình lưỡi liềm ở hai đầu đuôi, tương tự như cá mập và một số loài ichthyizard. Thay vì sử dụng các gợn sóng giống như rắn, cơ thể của chúng có thể vẫn cứng để giảm lực cản trong nước, trong khi đuôi của chúng tạo ra lực đẩy mạnh. [5] Những con vật này có thể ẩn nấp và vồ vập nhanh chóng và mạnh mẽ để vượt qua con mồi, thay vì đuổi theo nó. [6]
Việc tái tạo ban đầu cho thấy những con mosasaur với những chiếc mào lưng chạy dọc theo chiều dài cơ thể của chúng, dựa trên những xác nhận sai lầm của sụn khí quản. Vào thời điểm lỗi này được phát hiện, việc mô tả các mosasaur với các đỉnh như vậy trong tác phẩm nghệ thuật đã trở thành một xu hướng. [7][8]
Paleobiology [ chỉnh sửa ]




Mosasaur có hàm hai đầu và xương sọ linh hoạt rất giống với những con rắn), cho phép chúng nuốt chửng con mồi gần như toàn bộ. Một bộ xương của Tylosaurus proriger từ Nam Dakota bao gồm hài cốt của loài chim biển lặn Hesperornis một loài cá biển, một con cá mập có thể, và một con mosasaur nhỏ khác ( ). Xương Mosasaur cũng đã được tìm thấy với răng cá mập nhúng trong đó.
Một trong những món ăn của mosasaur là ammonite, động vật thân mềm có vỏ tương tự như Nautilus có rất nhiều ở vùng biển Creta. Các lỗ hổng đã được tìm thấy trong vỏ hóa thạch của một số loại ammonite, chủ yếu là Pachydiscus và Pl Nhauicera . Chúng từng được hiểu là kết quả của việc khập khiễng gắn vào ammonite, nhưng hình dạng tam giác của các lỗ, kích thước của chúng và sự hiện diện của chúng ở cả hai bên của vỏ, tương ứng với hàm trên và hàm dưới, là bằng chứng của vết cắn của môi trường mosasaurs kích thước. Cho dù hành vi này là phổ biến trên tất cả các lớp kích thước của mosasaur không rõ ràng.
Hầu như tất cả các dạng là động vật săn mồi tích cực của cá và ammonite; một số ít, chẳng hạn như Globidens có răng cùn, hình cầu, chuyên dùng để nghiền vỏ nhuyễn thể. Các chi nhỏ hơn, chẳng hạn như Platecarpus và Dallasaurus dài khoảng 1 đùa6 m (3,31919 ft), có thể được nuôi bằng cá và các con mồi nhỏ khác. Những con mosasaur nhỏ hơn có thể đã dành thời gian ở nước ngọt, săn lùng thức ăn. Các mosasaur lớn hơn, chẳng hạn như Tylosaurus Hainosaurus và Mosasaurus đạt kích thước dài 10 phút15 m (33 mối 49 ft) Các đại dương Creta muộn, tấn công các loài bò sát biển khác, cũng như săn bắt cá lớn và ammonite.
Mô mềm [ chỉnh sửa ]


Mặc dù vẫn còn nhiều kiến thức về thiên nhiên. của lớp phủ da của họ vẫn còn trong giai đoạn đầu của nó. Rất ít mẫu vật mosasaurid được thu thập từ khắp nơi trên thế giới lưu giữ những dấu ấn quy mô hóa thạch. Sự thiếu hụt này có thể là do tính chất tinh tế của vảy, gần như loại bỏ khả năng bảo quản, ngoài các loại trầm tích bảo quản và các điều kiện biển mà việc bảo quản xảy ra. Cho đến khi phát hiện ra một số mẫu vật mosasaur với các dấu ấn quy mô được bảo quản tốt đáng kể từ các mỏ tiền Maastrichtian của Muwaqqar Chalk Marl Formation of Mitchana [9] ở Jordan, kiến thức về bản chất của tích phân mosasaur chủ yếu dựa trên rất ít tài khoản mô tả có niên đại từ người Thượng Santonia hạ thấp, chẳng hạn như mẫu vật nổi tiếng Tylosaurus (KUVP-1075) từ Hạt Gove, Kansas. [10] Tài liệu từ Jordan đã cho thấy các thi thể của mosasaur, cũng như các màng giữa ngón tay và ngón chân của chúng, được bao phủ bởi các vảy nhỏ, chồng lên nhau, hình kim cương giống như của rắn. Giống như của các loài bò sát hiện đại, vảy mosasaur khác nhau trên cơ thể về chủng loại và kích cỡ. Trong các mẫu vật của Gurana, hai loại vảy được quan sát thấy trên một mẫu vật duy nhất: vảy nhọn bao phủ các vùng trên của cơ thể và vảy mịn bao phủ phần dưới. [9] Là kẻ săn mồi phục kích, ẩn nấp và nhanh chóng bắt giữ con mồi bằng chiến thuật tàng hình, [11] họ có thể đã được hưởng lợi từ các vảy không có tác dụng. [9]



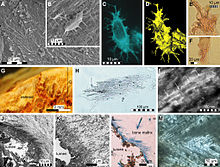
Vào năm 2011, protein collagen đã được phục hồi từ [194590020] humerus có niên đại kỷ Phấn trắng. [12]
Năm 2005, một nghiên cứu trường hợp của AS Schulp, E.W.A Mulder và K. Schwenk đã phác thảo thực tế rằng các mosasaur đã ghép đôi fenestrae trong vòm miệng của chúng. Trong thằn lằn và rắn theo dõi, fenestrae ghép nối được liên kết với lưỡi chĩa, được đưa vào và ra để phát hiện dấu vết hóa học và mang lại cảm giác về hướng. Do đó, họ đề xuất rằng mosasaur có lẽ cũng có lưỡi chẻ nhạy cảm. [13]
Trao đổi chất [ chỉnh sửa ]
Một nghiên cứu được công bố vào năm 2016 bởi T. Lyn Harrell, Alberto Pérez-Huerta và Celina Suarez cho thấy mosasaur là nhiệt nội. Nghiên cứu này mâu thuẫn với một nghiên cứu được công bố vào năm 2010 nói rằng mosasaur là sinh tinh. Nghiên cứu năm 2010 không sử dụng động vật máu nóng để so sánh nhưng các nhóm động vật biển phổ biến tương tự. Dựa trên sự so sánh với các động vật máu nóng hiện đại và hóa thạch của các động vật máu lạnh được biết đến trong cùng khoảng thời gian; nghiên cứu năm 2016 cho thấy những con mosasaur có thể có nhiệt độ cơ thể tương tự như loài chim biển hiện đại và có thể điều chỉnh bên trong nhiệt độ của chúng để giữ ấm hơn nước xung quanh. [14]
Coloration [ chỉnh sửa ]
Màu sắc của mosasaurs vẫn chưa được biết cho đến năm 2014, khi phát hiện của Johan Lindgren thuộc Đại học Lund và các đồng nghiệp đã tiết lộ sắc tố melanin trong vảy hóa thạch của một con mosasaur. Những con Mosasaur có khả năng là đầu nguồn, với lưng màu tối và lớp lông màu sáng, giống như một con cá mập trắng hoặc rùa biển da trắng, loài sau này có tổ tiên hóa thạch mà màu sắc cũng được xác định. Những phát hiện đã được mô tả trong tạp chí Thiên nhiên . [15]
Bản năng và sự tăng trưởng [ chỉnh sửa ]
Sự phát triển của Mosasaur không được hiểu rõ, vì mẫu vật của cá con rất hiếm, và nhiều người đã nhầm lẫn với loài chim ngần ngại khi được phát hiện 100 năm trước. Tuy nhiên, việc phát hiện ra một số mẫu vật của các con mosasaur ở tuổi vị thành niên và trẻ sơ sinh được khai quật cách đây hơn một thế kỷ cho thấy những con mosasaur đã sinh ra để sống và chúng sống những năm đầu đời ở ngoài biển, không phải trong các vườn ươm hoặc khu bảo tồn chẳng hạn như nước nông như trước đây tin tưởng. Nếu, giống như các loài bò sát biển khác (như plesiosaurs), mosasaur cung cấp sự chăm sóc của cha mẹ hiện chưa rõ. Việc phát hiện ra mosasaurs trẻ đã được công bố trên tạp chí Palaeontology . [16]
Môi trường [ chỉnh sửa ]
Các nhà cổ sinh vật học đã so sánh sự đa dạng về hình thái học và sự khác biệt về hình thái học. mực nước biển, nhiệt độ mặt nước biển và các đường cong đồng vị carbon ổn định cho kỷ Phấn trắng để khám phá các yếu tố có thể ảnh hưởng đến sự tiến hóa của chúng. Không có yếu tố duy nhất rõ ràng chiếm tất cả các bức xạ, đa dạng hóa và tuyệt chủng; tuy nhiên, các mô hình đa dạng hóa phân loại và hình thái học khác nhau rộng hơn dẫn đến sự khác biệt trong thị trường câu cá trên thuyền của Cameron dưới tác động của áp lực chọn lọc từ dưới lên trên. Động lực có khả năng nhất trong quá trình tiến hóa của mosasaur là năng suất cao trong kỷ Phấn trắng muộn, được thúc đẩy bởi mực nước biển được kiểm soát về mặt kiến tạo và phân tầng khí hậu và phân phối dinh dưỡng theo khí hậu. Khi năng suất sụp đổ vào cuối kỷ Phấn trắng, trùng hợp với tác động của bolide, mosasaur đã bị tuyệt chủng. [17]

Mực nước biển cao trong thời kỳ kỷ Phấn trắng, gây ra sự xâm phạm biển ở nhiều nơi trên thế giới và là một tuyến đường biển nội địa tuyệt vời. Bắc Mỹ. Hóa thạch Mosasaur đã được tìm thấy ở Hà Lan, Bỉ, Đan Mạch, Bồ Đào Nha, Thụy Điển, Nam Phi, Tây Ban Nha, Pháp, Đức, Ba Lan, Cộng hòa Séc, [18] Bulgaria, Vương quốc Anh, [19][20] Nga, Ukraine, Kazakhstan, Ailen, [21] Nhật Bản, [22] Ai Cập, Israel, Jordan, Syria, [23] Thổ Nhĩ Kỳ, [24] Nigeria, [25][26] Ăng-gô-la, Ma-rốc, Úc, New Zealand và trên đảo Vega ngoài khơi Nam Cực. Taxon răng Globidens timorensis được biết đến từ đảo Timor; tuy nhiên, vị trí phát sinh loài của loài này là không chắc chắn và nó thậm chí có thể không phải là một con mosasaur. [27] Mosasaur đã được tìm thấy ở Canada ở Manitoba và Saskatchewan [28] và ở hầu hết Hoa Kỳ. Các mẫu vật hoàn chỉnh hoặc một phần đã được tìm thấy ở Alabama, Mississippi, New Jersey, Tennessee và Georgia, cũng như ở các bang được bao phủ bởi đường biển Creta: Texas, tây nam Arkansas, New Mexico, Kansas, [29] Colorado, Nebraska, South Dakota , Montana, Wyoming, và các thành tạo Pierre Shale / Fox Hills ở Bắc Dakota. [30] Cuối cùng, xương và răng mosasaur cũng được biết đến từ California, Mexico, Colombia, [31] Brazil, [23] Peru và Chile. [32]
Nhiều người được gọi là "khủng long" được tìm thấy ở New Zealand thực sự là mosasaur và plesiosaur, cả hai đều là loài bò sát biển săn mồi Mesozoi.
Discovery [ chỉnh sửa ]


1764 bởi các công nhân mỏ đá trong một phòng trưng bày dưới mặt đất của một mỏ đá vôi ở Núi Saint Peter, gần thành phố Maastricht của Hà Lan, trước bất kỳ khám phá hóa thạch khủng long lớn nào, nhưng vẫn còn ít được biết đến. Tuy nhiên, một phát hiện thứ hai về hộp sọ một phần đã thu hút sự chú ý của Thời đại Khai sáng về sự tồn tại của các động vật hóa thạch khác với bất kỳ sinh vật sống nào được biết đến. Khi mẫu vật được phát hiện từ năm 1770 đến 1774, Johann Leonard Hoffmann, một nhà phẫu thuật và nhà sưu tập hóa thạch, đã trao đổi về nó với các nhà khoa học có ảnh hưởng nhất trong thời đại của ông, khiến hóa thạch trở nên nổi tiếng. Tuy nhiên, chủ sở hữu ban đầu là Godding, một giáo đường của nhà thờ Maastricht.
Khi các lực lượng cách mạng Pháp chiếm Maastricht vào năm 1794, hóa thạch được giấu cẩn thận đã được phát hiện, sau một phần thưởng, người ta nói, 600 chai rượu vang, và được vận chuyển đến Paris. Sau khi được giải thích trước đó là cá, cá sấu và cá nhà táng, người đầu tiên hiểu về mối quan hệ thằn lằn của nó là nhà khoa học người Hà Lan Adriaan Gilles Camper vào năm 1799. Năm 1808, Georges Cuvier đã xác nhận kết luận này, mặc dù le Grand Hóa thạch động vật de Maëstricht không thực sự được đặt tên là Mosasaurus ('Meuse reptile') cho đến năm 1822 và không được đặt tên đầy đủ của nó, Mosasaurus hoffmannii, cho đến năm 1829. hài cốt, đã được phát hiện trước đó tại Maastricht nhưng không được xác định là mosasaur cho đến thế kỷ 19, đã được trưng bày trong Bảo tàng Teylers, Haarlem, được mua từ năm 1790.
Những chiếc giường đá vôi Maastricht đã trở nên nổi tiếng nhờ phát hiện của mosasaur, chúng đã đặt tên cho kỷ nguyên sáu triệu năm cuối cùng của kỷ Phấn trắng, Maastrichtian.
Mối quan hệ [ chỉnh sửa ]
Bản sao của Mosasauridae được sửa đổi từ Simões et al. (2017): [33]
| Mosasauridae |
|
||||||
Lịch sử tiến hóa [ chỉnh sửa ]








Dựa trên các đặc điểm như hàng răng kép của răng cửa ("mặt bích") trên vòm miệng, hàm bị lỏng lẻo, hàm bị thay đổi Nhiều phương pháp xác định vị trí, nhiều nhà nghiên cứu tin rằng rắn có chung tổ tiên biển với mosasaur, một gợi ý được đưa ra vào năm 1869 bởi Edward Drinker, người đã đặt ra thuật ngữ Pythonomorpha để hợp nhất chúng. Ý tưởng nằm im lìm trong hơn một thế kỷ, được hồi sinh vào những năm 1990. [34][35] Gần đây, phát hiện Najash rionegrina một con rắn hóa thạch từ Nam Mỹ, nghi ngờ về giả thuyết nguồn gốc biển.
Bộ xương của Dallasaurus Turneri được mô tả bởi Bell và Polcyn (2005), có một hỗn hợp các đặc điểm hiện diện trong các bộ xương của mosasaur có nguồn gốc và trong bộ xương của tổ tiên mosasaurid, như aigialoid. Dallasaurus giữ lại các chi trên mặt đất tương tự về cấu trúc của chúng với các chi của aigializardids và squamate trên mặt đất (tình trạng chân tay bẩm sinh), không giống như các loại mosasaurids có nguồn gốc giống như mái chèo. Tuy nhiên, bộ xương của Dallasaurus đồng thời có một số nhân vật liên kết nó với các thành viên xuất phát của phân họ Mosasaurinae; các tác giả của mô tả của nó đã liệt kê "sự xâm lấn của paralal bởi các lưỡi trung gian từ phía trước, răng với bề mặt men trung gian mịn màng, bướu coronoid cao trên mũi khâu ngoài xương hàm, phẫu thuật cắt bỏ xương hàm, phẫu thuật cắt bỏ xương cụt synapophyses, các kiểu đốt sống định hướng theo chiều dọc, các đốt sống ngực kéo dài và các vòm cung hợp nhất "như các ký tự hợp nhất Dallasaurus với Mosasaurinae. [36] Phân tích phylogenetic. các nhánh không bao gồm các đơn vị phân loại, chẳng hạn như Dallasaurus Yaguaraaurus Russellosaurus Tethysaurus ] và Kiteriaaurus (năm 2005 chỉ được biết đến một cách không chính thức là "aigializard"); phân tích chỉ ra rằng tình trạng chân tay được phát triển độc lập ở ba nhóm mosasaur khác nhau (Halisaurinae, Mosasaurinae và nhóm có chứa các phân họ Tylizardinae và Plioplatecarpinae) đã được tiến hành bởi nghiên cứu này. Palci (2007) và Leblanc, Caldwell và Bardet (2012); [38][39] phân tích được thực hiện bởi Makádi, Caldwell và Ősi (2012) chỉ ra rằng tình trạng chân tay được tiến hóa độc lập ở hai nhóm mosasaurina và ở Mosasaurinae , Tylizardinae và Plioplatecarpinae). [40] Conrad và cộng sự (2011), mặt khác, các mosasaur đã thu hồi được tạo thành một dòng họ loại trừ họ hàng của họ. [41] Nếu giả thuyết của Bell và Polcyn (2005) là chính xác, thì mosasaur theo nghĩa truyền thống của từ này, nghĩa là "thằn lằn" rằng các chi giống như mái chèo tiến hóa và tỏa ra môi trường dưới nước vào cuối thời đại Trung sinh, bị tuyệt chủng vào cuối thời đại đó ", [37] thực sự là đa thê; Bell và Polcyn (2005) đã duy trì Mosasauridae độc quyền bằng cách bao gồm Dallasaurus và các loại taxi plaeopedal nói trên khác trong gia đình, [36] trong khi Caldwell (2012) đề nghị (mặc dù tuyên bố rõ ràng rằng đó không phải là "đề xuất chính thức" về danh pháp mới ") để hạn chế Mosasauridae chỉ trong chi Mosasaurus và họ hàng gần nhất của họ. [37]
Vị trí phát sinh loài chính xác của họ (aigializardids và dolichizards) trong Squamata vẫn chưa chắc chắn. Một số phân tích cladistic đã phục hồi chúng như là họ hàng gần nhất của rắn, [42][43] có tính đến sự tương đồng về giải phẫu hàm và sọ; [42] tuy nhiên, điều này đã bị tranh cãi [44][45][46] và phân tích hình thái học được thực hiện bởi Conrad (2008) Thay vào đó, varanoids liên quan chặt chẽ với thằn lằn giám sát mặt đất. [44] Phân tích tiếp theo về mối quan hệ anguimorph được thực hiện bởi Conrad và cộng sự (2011) dựa trên hình thái một mình đã thu hồi mosasaurids, aigializardids và dolichizards khi anguimorphs nằm bên ngoài các nhánh ít bao gồm có thằn lằn và helodermatids theo dõi; phân tích dựa trên các bộ dữ liệu kết hợp của dữ liệu hình thái và phân tử, mặt khác, cho thấy chúng có liên quan chặt chẽ hơn với thằn lằn giám sát và thằn lằn không tai so với helodermatids và thằn lằn cá sấu Trung Quốc. [41] Phân tích hình thái lớn được thực hiện bởi Gauthier và cộng sự (2012) đã thu hồi mosasaurids, aigializardids và dolichizardids ở vị trí không mong muốn là thành viên cơ bản của dòng họ Scincogekkonomorpha (chứa tất cả các loài taxi chia sẻ tổ tiên chung gần đây với Gekko gecko và Iguana iguana [44]) không thuộc về Scleroglossa. Vị trí phát sinh loài của các phân loại này hóa ra phụ thuộc rất nhiều vào việc phân loại hay đưa vào phân loại. Khi mosasaurids bị loại khỏi phân tích, dolichizards và aigializardids đã được phục hồi trong Scleroglossa, tạo thành một nhóm chị em với đội quân có rắn, amlawbaenian, dibamids và thằn lằn không chân Mỹ. Khi mosasaurids được đưa vào phân tích, và nhiều loại taxi khác nhau có các chi bị giảm hoặc vắng mặt ngoài rắn (như dibamids hoặc amlawbaenian) đã bị loại trừ, mosasaurids, aigializardids và dolichizard được thu hồi trong Scleroglossa để tạo thành nhóm chị em. Bhullar và Gauthier (2012) đã tiến hành phân tích hình thái các mối quan hệ squamate bằng cách sử dụng một phiên bản sửa đổi của ma trận từ phân tích của Gauthier et al. (2012); họ đã tìm thấy vị trí phát sinh loài của dòng họ chứa mosasaur và họ hàng gần nhất của chúng trong Squamata rất không ổn định, với dòng họ "được phục hồi bên ngoài Scleroglossa (như trong Gauthier và cộng sự 2012) các hình thức ". [48]
Phân phối [ chỉnh sửa ]
Mặc dù không có chi hoặc phân họ riêng lẻ nào được tìm thấy trên toàn thế giới, nhưng Mosasauridae đã đạt được sự phân phối toàn cầu trong kỷ Phấn trắng muộn với nhiều địa điểm thường phức tạp mosasaur faunas với nhiều chi và loài khác nhau trong các hốc sinh thái khác nhau.
Tài liệu tham khảo [ chỉnh sửa ]
- ^ Dash, Sean (2008). Quái vật thời tiền sử được tiết lộ . Hoa Kỳ: Kênh sản xuất / lịch sử tham công tiếc việc . Truy cập ngày 18 tháng 12, 2015 .
- ^ Lĩnh vực, Daniel J.; LeBlanc, Aaron; Gau1, Adrienne; Behlke, Adam D. (10 tháng 4 năm 2015). "Hóa thạch sơ sinh Pelagic hỗ trợ sinh lực và lịch sử cuộc sống tiền xã hội của các loài Hồi giáo Creta". Cổ sinh vật học . 58 (3): 401. doi: 10.1111 / pala.12165.
- ^ Grigoriev, D.W. (2014). "Khổng lồ Mosasaurus hoffmanni (Squamata, Mosasauridae) từ kỷ Phấn trắng muộn (Maastrichtian) của Penza, Nga" (PDF) . Thủ tục tố tụng của Viện Động vật học RAS . 318 (2): 148 Tiết167 . Truy cập 26 tháng 6 2016 .
- ^ "mosasaur lớn nhất được trưng bày". Kỷ lục Guinness thế giới . 2014 . Truy cập 27 tháng 6 2016 .
- ^ a b Lindgren, J.; Caldwell, M.W.; Konishi, T.; Chiappe, L.M. (2010). Farke, Andrew Allen, chủ biên. "Sự tiến hóa hội tụ trong Tetrapods dưới nước: Những hiểu biết sâu sắc về một Mosasaur hóa thạch đặc biệt". PLoS ONE . 5 (8): e11998. doi: 10.1371 / tạp chí.pone.0011998. PMC 2918493 . PMID 20711249.
- ^ Lindgren, J.; Kaddumi, H. F.; Polcyn, M. J. (2013). "Bảo tồn mô mềm trong một con thằn lằn biển hóa thạch với vây đuôi hai mắt". Truyền thông tự nhiên . 4 : 2423. doi: 10.1038 / ncomms3423. PMID 24022259.
- ^ Osborn, Henry Fairfield (1899). "Một bộ xương Mosasaur hoàn chỉnh, Osseous và sụn". Hồi ức của Bảo tàng Lịch sử Tự nhiên Hoa Kỳ . 1 (4): 167 Tiết188 . Truy cập 25 tháng 11 2014 .
- ^ Everhart, Mike (13 tháng 1 năm 2013). "Nguồn gốc của rìa lưng trên Mosasaur". Đại dương Kansas . Truy xuất ngày 25 tháng 11 2014 .
- ^ a b ] Kaddumi, HF (2009). "Trên các tấm phủ quy mô mới nhất của mosasaur (Squamata: Mosasauridae ) từ Động vật Mitchana ngoài mô tả về loài Mosasaurus mới". Hóa thạch của động vật Mitchana và các khu vực liền kề . Amman: Bảo tàng lịch sử tự nhiên sông vĩnh cửu. trang 80 bóng94.
- ^ Snow, F. H. (1878). "Trên lớp phủ của một loài bò sát mosasauroid". Giao dịch của Viện hàn lâm Khoa học Kansas . 6 : 54 Ảo58. doi: 10.2307 / 3623557. JSTOR 3623557.
- ^ Massare, J. A. (1987). "Hình thái răng và ưu tiên con mồi của các loài bò sát biển Mesozoi". Tạp chí Cổ sinh vật học Động vật có xương sống . 7 (2): 121 Chiếc137. doi: 10.1080 / 02724634.1987.10011647.
- ^ Lindgren, Johan; Uvdal, Per; Tiếng Anh, Tiếng Anh; Lee, Andrew H.; Alwmark, Carl; Bergquist, Karl-Erik; Nilsson, Einar; Kinh thánh, Peter; Rasmussen, Magnus; Douglas, Desirée A.; Polcyn, Michael J.; Jacobs, Louis L. (29 tháng 4 năm 2011). "Bằng chứng hiển vi của các protein xương Creta". PLoS ONE . 6 (4): e19445. doi: 10.1371 / tạp chí.pone.0019445. ISSN 1932-6203. PMC 3084868 . PMID 21559386.
- ^ Schulp, A. S.; Mulder, E. W. A.; Schwenk, K. (2005-09-01). "Có phải mosasaur đã chĩa lưỡi?". Tạp chí khoa học địa chất Hà Lan . 84 (3): 359 Điêu371. doi: 10.1017 / S0016774600021144. ISSN 0016-7746 – thông qua Nhà xuất bản Đại học Cambridge.
- ^ "Các nhà khoa học trích dẫn bằng chứng cho thấy mosasaur có máu nóng". Khoa học hàng ngày . Đã truy xuất 2017-08-01 .
- ^ Lindgren, J.; Sjövall, P.; Carney, R. M.; Uvdal, P.; Gren, J. A.; Đê, G.; Schultz, B. P.; Khăn choàng, M. D.; Barnes, K. R.; Polcyn, M. J. (2014). "Sắc tố da cung cấp bằng chứng về melanism hội tụ trong các loài bò sát biển đã tuyệt chủng". Thiên nhiên . 506 (7361): 484 Tắt8. doi: 10.1038 / thiên nhiên12899. PMID 24402224.
- ^ "Cuộc sống của những con thằn lằn biển khổng lồ mới sinh trong thời đại khủng long". Khoa học hàng ngày . Đã truy xuất 2017-08-01 .
- ^ Polcyn, M. J.; Jacobs, L.; Araujo, R.; Schulp, A. S.; Mateus, O. (2014). "Trình điều khiển vật lý của sự tiến hóa mosasaur". Palaeogeography, Palaeoclimatology, Palaeoecology . 400 : 17 Hàng27. doi: 10.1016 / j.palaeo.2013.05.018.
- ^ "Druhohorní plazi v Čechách II". DinosaurusBlog . 2015/07/13 . Truy xuất 2017-08-01 .
- ^ "St James 'Pit, Norwich (SSSI)" (PDF) . Nước Anh tự nhiên . 2014 . Truy cập 25 tháng 11 2014 .
- ^ Jagt, John W. M.; Motchurova-Dekova, Neda; Ivanov, Plamen; Cappetta, Hen-ri; Schulp, Anne S. (2006). "Những con thằn lằn mới nhất và cá mập lamniform từ hang Labirinta, quận Vratsa (tây bắc Bulgaria): Một ghi chú sơ bộ". Geoloski Anali Balkanskoga Poluostrva . 67 (67): 51 Tái63. doi: 10.2298 / gabp0667051j.
- ^ Storrs, Glenn W.; Arkhangelskii, Maxim S.; Efimov, Vladimir M. (2000). "Các loài bò sát biển Mesozoi của Nga và các nước cộng hòa thuộc Liên Xô cũ khác". Ở Benton, M. J.; Shishkin, M. A.; Unwin, D. M. Thời đại khủng long ở Nga và Mông Cổ . Cambridge: Nhà xuất bản Đại học Cambridge. trang 187 mỏ210. SĐT 980-0521554763.
- ^ Konishi, Takuya; Tanimoto, Masahiro; Utsunomiya, Satoshi; Sato, Masahiro; Watanabe, Katsunori (2012). "Một loài Mosasaurine lớn (Squamata: Mosasauridae) từ kỷ Phấn trắng mới nhất của tỉnh Osaka (Sw Nhật Bản)". Nghiên cứu cổ sinh vật học . 16 (2): 79 Kiếm87. doi: 10.2517 / 1342-8144-16.2.079.
- ^ a b Bardet, Nathalie; Pereda Suberbiola, Xabier; Iarochène, Mohamed; Amalik, Mohamed; Bouya, Baadi (tháng 9 năm 2005). "Durophagous Mosasauridae (Squamata) từ Phosphate thượng lưu của Morocco, với mô tả về một loài mới của Globidens ". Tạp chí khoa học địa chất Hà Lan . 84 (3): 167 Linh175. doi: 10.1017 / S0016774600020953. ISSN 0016-7746 – thông qua Nhà xuất bản Đại học Cambridge.
- ^ Bardet, Nathalie; Tunoğlu, Cemal (19 tháng 9 năm 2002) [24 Aug 2010]. "Mosasaur đầu tiên (Squamata) từ kỷ Phấn trắng muộn của Thổ Nhĩ Kỳ". Tạp chí Cổ sinh vật học Động vật có xương sống . 22 (3): 712 Ảo715. doi: 10.1671 / 0272-4634 (2002) 022 [0712:TFMSFT] 2.0.CO; 2. ISSN 0272-4634.
- ^ Lingham-Soliar, Theagarten (1991). "Mosasaurs từ kỷ Phấn trắng trên của Nigeria". Cổ sinh vật học . 34 (3): 653 Ảo670 – thông qua BioStor. [ liên kết chết vĩnh viễn ]
- ^ Lingham-Soliar, Theagarten (1998). "Một con mosasaur mới Pluridens walkeri từ Thượng Creta, Maastrichtian của Lưu vực Iullemmeden, phía tây nam Nigeria". Tạp chí Cổ sinh vật học Động vật có xương sống . 18 (4): 709 Ảo717. doi: 10.1080 / 02724634.1998.10011100.
- ^ Martin, James E. (2007). "Một loài mới của loài muỗi khổng lồ, Globidens (Squamata: Mosasauridae) từ Tập đoàn Pierre Shale muộn của miền trung Nam Dakota, Hoa Kỳ". Ở Martin, James E.; Parris, David C. Địa chất và Cổ sinh vật học của các trầm tích biển kỷ Phấn trắng muộn của Dakotas . 427 . Hiệp hội Địa chất Hoa Kỳ. tr 177 1771919. doi: 10.1130 / 2007.2427 (13). Sê-ri 980-0-8137-2427-0.
- ^ "Thông tin chung". Trung tâm khám phá hóa thạch Canada . 2014 . Truy cập 25 tháng 11 2014 .
- ^ Michael J. Everhart (2005). "Chương 9: Nhập Mosasaur". Đại dương Kansas: lịch sử tự nhiên của biển nội địa phía tây . Bloomington: Nhà xuất bản Đại học Indiana. Sê-ri 980-0-253-34547-9.
- ^ Getman, Myron (1994). Sự xuất hiện của Mosasaur và các hóa thạch bò sát khác từ hệ tầng Fox Hills (Maastrichtian: cuối kỷ Phấn trắng) của Bắc Dakota (Luận án danh dự địa chất). Đại học St. Lawrence thuộc Khoa Địa chất.
- ^ Páramo-Fonseca, María Eurídice (1 tháng 3 năm 2012). "Mosasauroids từ Colombia". Bulletin de la Société Géologique de France . 183 (2): 103 Tái 109. doi: 10.2113 / gssgfbull.183.2.103. ISSN 0037-9409 – thông qua GeoScienceWorld.
- ^ Otero, Rodrigo A.; Parham, James F.; Soto-Acuña, Sergio; Jimenez-Huidobro, Paulina; Rubilar-Rogers, David (2012). "Marine reptiles from Late Cretaceous (early Maastrichtian) deposits in Algarrobo, central Chile". Cretaceous Research. 35: 124–132. doi:10.1016/j.cretres.2011.12.003.
- ^ Simões, Tiago R.; Vernygora, Oksana; Paparella, Ilaria; Jimenez-Huidobro, Paulina; Caldwell, Michael W. (2017-05-03). "Mosasauroid phylogeny under multiple phylogenetic methods provides new insights on the evolution of aquatic adaptations in the group". PLOS ONE. 12 (5): e0176773. doi:10.1371/journal.pone.0176773. ISSN 1932-6203. PMID 28467456.
- ^ "Palaeos Vertebrates Squamata: Pythonomorpha". palaeos.com. 2012. Retrieved 25 November 2014.
- ^ Everhart, M. J. (2000). "Mosasaurs: Last of the Great Marine Reptiles". Prehistoric Times (44): 29–31. Retrieved 25 November 2014.
- ^ a b c Bell, G. L.; Polcyn, M. J. (Sep 2005). "Dallasaurus turneria new primitive mosasauroid from the Middle Turonian of Texas and comments on the phylogeny of Mosasauridae (Squamata)". Netherlands Journal of Geosciences. 84 (3): 177–194. doi:10.1017/S0016774600020965. ISSN 0016-7746 – via ResearchGate.
- ^ a b c Caldwell, Michael W. (2012-01-01). "A challenge to categories: "What, if anything, is a mosasaur?"". Bulletin de la Société Géologique de France. 183 (1): 7–34. doi:10.2113/gssgfbull.183.1.7. ISSN 0037-9409.
- ^ Leblanc, Aaron R. H.; Caldwell, Michael W.; Bardet, Nathalie (January 2012). "A new mosasaurine from the Maastrichtian (Upper Cretaceous) phosphates of Morocco and its implications for mosasaurine systematics". Journal of Vertebrate Paleontology. 32 (1): 82–104. doi:10.1080/02724634.2012.624145. ISSN 0272-4634.
- ^ Caldwell, Michael W.; Palci, Alessandro (2007-12-12). "A new basal mosasauroid from the Cenomanian (U. Cretaceous) of Slovenia with a review of mosasauroid phylogeny and evolution". Journal of Vertebrate Paleontology. 27 (4): 863–880. doi:10.1671/0272-4634(2007)27[863:ANBMFT]2.0.CO;2. ISSN 0272-4634.
- ^ Makádi, L. S.; Caldwell, M. W.; Ősi, A. (2012) [12 Jan 2012]. Butler, Richard J, ed. "The First Freshwater Mosasauroid (Upper Cretaceous, Hungary) and a New Clade of Basal Mosasauroids". PLoS ONE . 7 (12): e51781. doi:10.1371/journal.pone.0051781. PMC 3526648. PMID 23284766.
- ^ a b Conrad, Jack L.; Ast, Jennifer C.; Montanari, Shaena; Norell, Mark A. (22 July 2010). "A combined evidence phylogenetic analysis of Anguimorpha (Reptilia: Squamata)". Cladistics. 27 (3): 230–277. doi:10.1111/j.1096-0031.2010.00330.x. ISSN 0748-3007.
- ^ a b Lee, Michael S. Y. (29 January 1997). "The phylogeny of varanoid lizards and the affinities of snakes". Philosophical Transactions of the Royal Society of London B: Biological Sciences. 352 (1349): 53–91. doi:10.1098/rstb.1997.0005. ISSN 0962-8436. PMC 1691912.
- ^ Lee, Michael S. Y. (22 June 2005). "Molecular evidence and marine snake origins". Biology Letters. 1 (2): 227–230. doi:10.1098/rsbl.2004.0282. ISSN 1744-9561. PMC 1626205. PMID 17148173.
- ^ a b c Conrad, Jack L. (3 June 2008). "Phylogeny And Systematics Of Squamata (Reptilia) Based On Morphology". Bulletin of the American Museum of Natural History. 310: 1–182. doi:10.1206/310.1. ISSN 0003-0090.
- ^ Vidal, Nicolas; Hedges, S. Blair (7 May 2004). "Molecular evidence for a terrestrial origin of snakes". Proceedings of the Royal Society of London B: Biological Sciences. 271 (Suppl 4): S226–S229. doi:10.1098/rsbl.2003.0151. ISSN 0962-8452. PMC 1810015. PMID 15252991.
- ^ Apesteguía, Sebastián; Zaher, Hussam (April 2006). "A Cretaceous terrestrial snake with robust hindlimbs and a sacrum". Thiên nhiên . 440 (7087): 1037–1040. doi:10.1038/nature04413. ISSN 0028-0836. PMID 16625194.
- ^ Gauthier, Jacques A.; Kearney, Maureen; Maisano, Jessica Anderson; Rieppel, Olivier; Behlke, Adam D.B. (April 2012). "Assembling the Squamate Tree of Life: Perspectives from the Phenotype and the Fossil Record". Bulletin of the Peabody Museum of Natural History. 53 (1): 3–308. doi:10.3374/014.053.0101. ISSN 0079-032X.
- ^ Longrich, Nicholas R.; Bhullar, Bhart-Anjan S.; Gauthier, Jacques A. (26 December 2012). "Mass extinction of lizards and snakes at the Cretaceous–Paleogene boundary". Proceedings of the National Academy of Sciences. 109 (52): 21396–21401. doi:10.1073/pnas.1211526110. ISSN 0027-8424. PMC 3535637. PMID 23236177.
External links[edit]