Sinh học phát triển là nghiên cứu về quá trình động vật và thực vật sinh trưởng và phát triển. Sinh học phát triển cũng bao gồm sinh học tái sinh, sinh sản vô tính, biến thái và sự phát triển và biệt hóa của các tế bào gốc trong cơ thể người trưởng thành.
Vào cuối thế kỷ 20, ngành học phần lớn chuyển thành sinh học phát triển tiến hóa.
Quan điểm [ chỉnh sửa ]
Các quá trình chính liên quan đến sự phát triển phôi của động vật là: đặc tả khu vực, hình thái học, phân biệt tế bào, tăng trưởng và kiểm soát tổng thể thời gian được khám phá trong quá trình tiến hóa sinh học phát triển:
- Đặc tả khu vực đề cập đến các quá trình tạo ra mô hình không gian trong một quả bóng hoặc một tấm của các ô tương tự ban đầu. Điều này thường liên quan đến hoạt động của các yếu tố quyết định tế bào chất, nằm trong các bộ phận của trứng được thụ tinh và các tín hiệu cảm ứng phát ra từ các trung tâm tín hiệu trong phôi. Các giai đoạn đầu của đặc tả khu vực không tạo ra các tế bào biệt hóa chức năng, nhưng các quần thể tế bào cam kết phát triển đến một khu vực cụ thể hoặc một phần của sinh vật. Chúng được xác định bởi sự biểu hiện của sự kết hợp cụ thể của các yếu tố phiên mã.
- Morphogenesis liên quan đến sự hình thành hình dạng ba chiều. Nó chủ yếu liên quan đến các chuyển động được phối hợp của các tấm tế bào và của các tế bào riêng lẻ. Morphogenesis rất quan trọng để tạo ra ba lớp mầm của phôi sớm (ectoderm, mesoderm và endoderm) và để xây dựng các cấu trúc phức tạp trong quá trình phát triển cơ quan.
- Sự biệt hóa tế bào liên quan cụ thể đến sự hình thành các loại tế bào chức năng như thần kinh, cơ bắp, biểu mô tiết, vv Các tế bào biệt hóa chứa một lượng lớn protein cụ thể liên quan đến chức năng tế bào.
- Tăng trưởng bao gồm cả sự tăng kích thước tổng thể, và cả sự tăng trưởng khác biệt của các bộ phận (allometry) góp phần hình thành. Sự tăng trưởng chủ yếu xảy ra thông qua sự phân chia tế bào mà còn thông qua sự thay đổi kích thước tế bào và sự lắng đọng của các vật liệu ngoại bào. Hiện vẫn chưa rõ liệu phôi động vật có cơ chế đồng hồ chủ hay không.
Sự phát triển của thực vật bao gồm các quá trình tương tự như của động vật. Tuy nhiên, tế bào thực vật chủ yếu là bất động nên sự phát sinh hình thái đạt được bằng sự tăng trưởng khác biệt, không có sự di chuyển của tế bào. Ngoài ra, các tín hiệu quy nạp và các gen liên quan khác với các tín hiệu điều khiển sự phát triển của động vật.
Các quá trình phát triển [ chỉnh sửa ]
Phân biệt tế bào [ chỉnh sửa ]
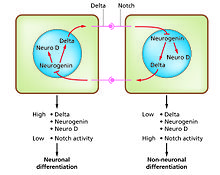
Phân biệt tế bào là quá trình phát sinh các loại tế bào chức năng khác nhau. Ví dụ, tế bào thần kinh, sợi cơ và tế bào gan (tế bào gan) là những loại tế bào biệt hóa nổi tiếng. Các tế bào biệt hóa thường tạo ra một lượng lớn một vài protein cần thiết cho chức năng cụ thể của chúng và điều này mang lại cho chúng vẻ ngoài đặc trưng cho phép chúng được nhận dạng dưới kính hiển vi ánh sáng. Các gen mã hóa các protein này có hoạt tính cao. Thông thường, cấu trúc nhiễm sắc thể của chúng rất mở, cho phép truy cập các enzyme phiên mã và các yếu tố phiên mã cụ thể liên kết với các chuỗi điều hòa trong DNA để kích hoạt biểu hiện gen. [1][2] Ví dụ, NeuroD là yếu tố phiên mã quan trọng cho sự biệt hóa tế bào thần kinh, myogenin cho sự biệt hóa cơ bắp, và HNF4 cho sự biệt hóa tế bào gan.
Sự biệt hóa tế bào thường là giai đoạn phát triển cuối cùng, trước một số trạng thái cam kết không được phân biệt rõ ràng. Một mô duy nhất, được hình thành từ một loại tế bào tiền thân hoặc tế bào gốc, thường bao gồm một số loại tế bào khác biệt. Kiểm soát sự hình thành của chúng liên quan đến quá trình ức chế bên, [3] dựa trên các đặc tính của đường dẫn tín hiệu Notch. [4] Ví dụ, trong tấm thần kinh của phôi, hệ thống này hoạt động để tạo ra một quần thể tế bào tiền thân thần kinh, trong đó NeuroD được thể hiện cao.
Tái sinh [ chỉnh sửa ]
Tái sinh cho thấy khả năng tái sinh một phần bị thiếu. [5] Điều này rất phổ biến ở các loài thực vật, cho thấy sự phát triển liên tục và cả trong số các động vật thuộc địa như vậy. như hydroids và ascidians. Nhưng hầu hết sự quan tâm của các nhà sinh học phát triển đã được thể hiện trong việc tái tạo các bộ phận ở động vật sống tự do. Đặc biệt bốn mô hình đã là chủ đề của nhiều cuộc điều tra. Hai trong số này có khả năng tái tạo toàn bộ cơ thể: Hydra có thể tái tạo bất kỳ phần nào của polyp từ một mảnh nhỏ, [6] và giun hành tinh, thường có thể tái tạo cả đầu và đuôi. [19659027] Cả hai ví dụ này đều có sự thay đổi tế bào liên tục được nuôi dưỡng bởi các tế bào gốc và, ít nhất là ở Planaria, ít nhất một số tế bào gốc đã được chứng minh là đa năng. [8] Hai mô hình còn lại chỉ cho thấy sự tái tạo của các phần phụ. Đây là các phần phụ của côn trùng, thường là chân của các loài côn trùng hemimetabolous như dế, [9] và tứ chi của động vật lưỡng cư urodele. [10] Thông tin đáng chú ý hiện có sẵn về sự tái sinh của chi lưỡng cư và được biết rằng mỗi loại tế bào đều tự tái sinh. ngoại trừ các mô liên kết nơi có sự giao thoa đáng kể giữa sụn, hạ bì và gân. Về mặt mô hình cấu trúc, điều này được kiểm soát bằng cách kích hoạt lại các tín hiệu hoạt động trong phôi. Vẫn còn nhiều tranh cãi về câu hỏi cũ về việc tái sinh là một tài sản "nguyên sơ" hay "thích nghi". [11] Nếu trước đây, với kiến thức được cải thiện, chúng ta có thể hy vọng có thể cải thiện khả năng tái sinh ở người. Nếu sau này, thì mỗi trường hợp tái sinh được cho là đã phát sinh do chọn lọc tự nhiên trong hoàn cảnh cụ thể của loài, do đó, không có quy tắc chung nào được mong đợi.
Sự phát triển phôi của động vật [ chỉnh sửa ]
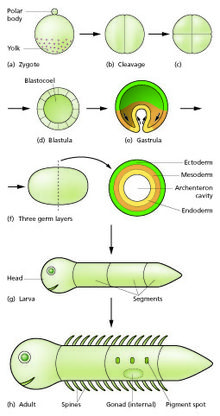

Cầu chì tinh trùng và trứng trong quá trình thụ tinh để hình thành trứng được thụ tinh, hoặc hợp tử. [12] Điều này trải qua một giai đoạn phân chia để tạo thành một quả bóng hoặc các tế bào tương tự được gọi là bóng một blastula hoặc blastoderm. Các phân chia tế bào này thường nhanh chóng và không có sự phát triển nên các tế bào con có kích thước bằng một nửa tế bào mẹ và toàn bộ phôi vẫn giữ nguyên kích thước. Chúng được gọi là sự phân chia. Các chuyển động hình thái chuyển đổi khối tế bào thành một cấu trúc ba lớp bao gồm các tấm đa bào gọi là ectoderm, mesoderm và endoderm, được gọi là các lớp mầm. Đây là quá trình của bệnh dạ dày. Trong quá trình phân tách và tập trung, các sự kiện đặc tả khu vực đầu tiên xảy ra. Ngoài việc hình thành ba lớp mầm, chúng thường tạo ra các cấu trúc ngoại bào, như nhau thai động vật, cần thiết cho sự hỗ trợ và dinh dưỡng của phôi, [13] và cũng thiết lập sự khác biệt về cam kết dọc theo trục trước (đầu, thân và đuôi). [14]
Đặc điểm kỹ thuật khu vực được bắt đầu bởi sự hiện diện của các yếu tố quyết định tế bào chất trong một phần của hợp tử. Các tế bào chứa yếu tố quyết định trở thành một trung tâm tín hiệu và phát ra một yếu tố cảm ứng. Bởi vì yếu tố cảm ứng được tạo ra ở một nơi, khuếch tán và phân rã, nó tạo thành một dải nồng độ, cao gần các tế bào nguồn và thấp hơn nữa. [15][16] Các tế bào còn lại của phôi, không chứa yếu tố quyết định, là có khả năng đáp ứng với các nồng độ khác nhau bằng cách điều chỉnh các gen kiểm soát phát triển cụ thể. Điều này dẫn đến một loạt các khu vực được thiết lập, được sắp xếp ở khoảng cách dần dần từ trung tâm tín hiệu. Ở mỗi khu vực, một tổ hợp gen kiểm soát phát triển khác nhau được điều hòa. [17] Những gen này mã hóa các yếu tố phiên mã điều chỉnh các tổ hợp hoạt động gen mới ở mỗi khu vực. Trong số các chức năng khác, các yếu tố phiên mã này kiểm soát sự biểu hiện của các gen tạo ra các đặc tính kết dính và vận động cụ thể trên các tế bào mà chúng hoạt động. Do các đặc tính hình thái khác nhau này, các tế bào của mỗi lớp mầm di chuyển tạo thành các tấm sao cho lớp ngoài tử cung ở bên ngoài, trung bì ở giữa và nội nhũ ở bên trong. [18][19] Chuyển động hình thái không chỉ thay đổi hình dạng và cấu trúc. của phôi, nhưng bằng cách đưa các tấm tế bào vào các mối quan hệ không gian mới, chúng cũng tạo ra các giai đoạn mới của tín hiệu và phản ứng giữa chúng.
Sự tăng trưởng trong phôi chủ yếu là tự trị. [20] Đối với mỗi lãnh thổ của tế bào, tốc độ tăng trưởng được kiểm soát bởi sự kết hợp của các gen đang hoạt động. Phôi sống tự do không phát triển hàng loạt vì chúng không có nguồn cung cấp thực phẩm bên ngoài. Nhưng phôi được cung cấp bởi nhau thai hoặc cung cấp lòng đỏ ngoài cơ thể có thể phát triển rất nhanh, và thay đổi tốc độ tăng trưởng tương đối giữa các bộ phận trong các sinh vật này giúp tạo ra giải phẫu tổng thể cuối cùng.
Toàn bộ quá trình cần được phối hợp kịp thời và cách thức kiểm soát điều này không được hiểu rõ. Có thể có một chiếc đồng hồ chủ có thể giao tiếp với tất cả các bộ phận của phôi điều khiển tiến trình của sự kiện, hoặc thời gian có thể phụ thuộc đơn giản vào chuỗi sự kiện nguyên nhân cục bộ của sự kiện. [21]
Biến thái [ chỉnh sửa ]
Các quá trình phát triển rất rõ ràng trong quá trình biến thái. Điều này xảy ra trong các loại động vật. Nổi tiếng là những ví dụ của ếch, thường nở ra như một con nòng nọc và biến chất thành ếch trưởng thành, và một số côn trùng nhất định nở thành ấu trùng và sau đó được tái tạo thành hình dạng trưởng thành trong giai đoạn nhộng.
Tất cả các quá trình phát triển được liệt kê ở trên xảy ra trong quá trình biến chất. Các ví dụ đã được nghiên cứu đặc biệt bao gồm mất đuôi và những thay đổi khác trong nòng nọc của ếch Xenopus [22][23] và sinh học của các đĩa tưởng tượng, tạo ra các bộ phận cơ thể trưởng thành của ruồi Drosophila melanogaster . [24][25]
Phát triển thực vật [ chỉnh sửa ]
Phát triển thực vật là quá trình mà các cấu trúc bắt nguồn và trưởng thành khi cây phát triển. Nó được nghiên cứu về giải phẫu thực vật và sinh lý thực vật cũng như hình thái thực vật.
Thực vật liên tục tạo ra các mô và cấu trúc mới trong suốt cuộc đời của chúng từ mô phân sinh [26] nằm ở đầu các cơ quan, hoặc giữa các mô trưởng thành. Do đó, một cây sống luôn có các mô phôi thai. Ngược lại, một phôi động vật sẽ sớm sản xuất tất cả các bộ phận cơ thể mà nó sẽ có trong cuộc sống của nó. Khi con vật được sinh ra (hoặc nở ra từ trứng của nó), nó có tất cả các bộ phận cơ thể và từ đó sẽ chỉ lớn hơn và trưởng thành hơn.
Các thuộc tính của tổ chức nhìn thấy trong một nhà máy là các thuộc tính nổi bật nhiều hơn tổng của các bộ phận riêng lẻ. "Việc tập hợp các mô và chức năng này thành một sinh vật đa bào tích hợp mang lại không chỉ các đặc điểm của các bộ phận và quá trình riêng biệt mà còn có một loạt các đặc điểm mới không thể dự đoán được trên cơ sở kiểm tra các bộ phận riêng biệt." [19659056] Tăng trưởng [ chỉnh sửa ]
Một thực vật có mạch bắt đầu từ một hợp tử đơn bào, được hình thành bằng cách thụ tinh của một tế bào trứng bởi một tế bào tinh trùng. Từ thời điểm đó, nó bắt đầu phân chia để hình thành phôi thực vật thông qua quá trình tạo phôi. Khi điều này xảy ra, các ô kết quả sẽ tổ chức sao cho một đầu trở thành gốc đầu tiên, trong khi đầu kia tạo thành đỉnh của chồi. Trong các nhà máy hạt giống, phôi sẽ phát triển một hoặc nhiều "lá giống" (lá mầm). Khi kết thúc quá trình tạo phôi, cây non sẽ có tất cả các bộ phận cần thiết để bắt đầu trong cuộc sống của nó.
Sau khi phôi nảy mầm từ hạt giống hoặc cây mẹ, nó bắt đầu sản xuất thêm các cơ quan (lá, thân và rễ) thông qua quá trình phát sinh cơ quan. Rễ mới mọc ra từ mô phân sinh rễ ở đầu rễ, thân và lá mới mọc ra từ mô phân sinh nằm ở đầu chồi. [28] Sự phân nhánh xảy ra khi các cụm tế bào nhỏ bị bỏ lại bởi mô phân sinh và có chưa trải qua quá trình biệt hóa tế bào để tạo thành một mô chuyên biệt, bắt đầu phát triển như là đỉnh của một rễ hoặc chồi mới. Tăng trưởng từ bất kỳ mô phân sinh nào ở đầu rễ hoặc chồi được gọi là tăng trưởng chính và dẫn đến việc kéo dài gốc hoặc chồi đó. Sự tăng trưởng thứ cấp dẫn đến việc mở rộng một rễ hoặc bắn ra từ sự phân chia của các tế bào trong một cambium. [29]
Ngoài sự tăng trưởng bằng cách phân chia tế bào, một cây có thể phát triển qua sự kéo dài tế bào ]. Điều này xảy ra khi các tế bào riêng lẻ hoặc nhóm các tế bào phát triển dài hơn. Không phải tất cả các tế bào thực vật sẽ phát triển đến cùng một chiều dài. Khi các tế bào ở một bên của thân phát triển dài hơn và nhanh hơn các tế bào ở phía bên kia, kết quả là thân cây sẽ uốn cong sang một bên của các tế bào phát triển chậm hơn. Sự tăng trưởng định hướng này có thể xảy ra thông qua phản ứng của thực vật đối với một kích thích cụ thể, chẳng hạn như ánh sáng (phototropism), trọng lực (hấp dẫn), nước, (hydrotropism) và tiếp xúc vật lý (thigmotropism).
Sự tăng trưởng và phát triển của thực vật được điều hòa bởi các hoocmon và chất điều hòa sinh trưởng thực vật cụ thể (PGRs) (Ross và cộng sự 1983). [30] Nồng độ hormone nội sinh bị ảnh hưởng bởi tuổi cây, độ cứng lạnh, ngủ đông và các điều kiện trao đổi chất khác; photoperiod, hạn hán, nhiệt độ và các điều kiện môi trường bên ngoài khác; và các nguồn PGR ngoại sinh, ví dụ, được áp dụng bên ngoài và có nguồn gốc rhizospheric.
Biến đổi hình thái [ chỉnh sửa ]
Thực vật thể hiện sự biến đổi tự nhiên trong hình dạng và cấu trúc của chúng. Trong khi tất cả các sinh vật khác nhau từ cá nhân đến cá nhân, thực vật thể hiện một loại biến thể bổ sung. Trong một cá nhân, các bộ phận được lặp lại có thể khác nhau về hình thức và cấu trúc so với các bộ phận tương tự khác. Sự biến đổi này dễ thấy nhất ở lá của cây, mặc dù các cơ quan khác như thân và hoa có thể cho thấy sự biến đổi tương tự. Có ba nguyên nhân chính của sự biến đổi này: hiệu ứng vị trí, tác động môi trường và tuổi vị thành niên.
Sự tiến hóa của hình thái thực vật [ chỉnh sửa ]
Các yếu tố phiên mã và mạng lưới điều hòa phiên mã đóng vai trò quan trọng trong hình thái thực vật và sự tiến hóa của chúng. Trong quá trình hạ cánh thực vật, nhiều họ yếu tố phiên mã mới xuất hiện và được ưu tiên kết nối với các mạng lưới phát triển đa bào, sinh sản và phát triển cơ quan, góp phần hình thành các hình thái phức tạp hơn của thực vật trên đất. [31]
Sinh vật mô hình phát triển ]
Phần lớn nghiên cứu sinh học phát triển trong những thập kỷ gần đây đã tập trung vào việc sử dụng một số lượng nhỏ các sinh vật mô hình. Nó đã chỉ ra rằng có nhiều bảo tồn các cơ chế phát triển trên khắp vương quốc động vật. Trong giai đoạn đầu phát triển, các loài động vật có xương sống khác nhau, tất cả đều sử dụng các tín hiệu quy nạp giống nhau và cùng các gen mã hóa bản sắc khu vực. Ngay cả động vật không xương sống cũng sử dụng một tiết mục tương tự về tín hiệu và gen mặc dù các bộ phận cơ thể được hình thành khác nhau đáng kể. Mỗi sinh vật mẫu có một số lợi thế thí nghiệm đặc biệt cho phép chúng trở nên phổ biến trong các nhà nghiên cứu. Theo một nghĩa nào đó, chúng là "mô hình" cho toàn bộ vương quốc động vật và theo một nghĩa khác, chúng là "mô hình" cho sự phát triển của con người, rất khó để nghiên cứu trực tiếp vì cả lý do đạo đức và thực tế. Các sinh vật mẫu đã hữu ích nhất để làm sáng tỏ bản chất rộng lớn của các cơ chế phát triển. Càng tìm kiếm nhiều chi tiết, chúng càng khác biệt với nhau và với con người.
Thực vật:
Động vật có xương sống:
- Ếch: Xenopus ( X.laevis và nhiệt đới ). [32][33] Cung cấp phôi tốt. Đặc biệt thích hợp cho ngành vi phẫu.
- Zebrafish: Danio rerio . [34] Cung cấp phôi tốt. Di truyền học phát triển tốt.
- Gà: Gallus gallus . [35] Giai đoạn đầu tương tự như động vật có vú, nhưng vi phẫu dễ dàng hơn. Chi phí thấp.
- Chuột: Mus musculus . [36] Một động vật có vú có di truyền phát triển tốt.
Động vật không xương sống:
Cũng phổ biến cho một số mục đích là nhím biển
và ascidians. [40] Đối với các nghiên cứu về động vật lưỡng cư tái sinh urodele như axolotl Ambystoma mexicanum được sử dụng, [41] và cả các loại giun hành tinh như Schmidtea cũng được chứng minh là một mô hình hiệu quả cho sự phát triển. [43] Sự phát triển của thực vật đã tập trung vào cây cải xoong Arabidopsis thaliana với tư cách là một sinh vật mẫu. [44]
Xem thêm [ chỉnh sửa ]
Tài liệu tham khảo [ chỉnh sửa ]
- ^ Li B.; Cẩn thận M.; Công nhân J.L. (2007). "Vai trò của chất nhiễm sắc trong quá trình phiên mã". Tế bào . 128 : 707 Từ719. doi: 10.1016 / j.cell.2007.01.015. PMID 17320508.
- ^ Heintzman N.D.; et al. (2007). "Chữ ký chromatin khác biệt và dự đoán của các chất thúc đẩy phiên mã và chất tăng cường trong bộ gen của con người". Nat Genet . 39 : 311 trừ318. doi: 10.1038 / ng1966. PMID 17277777.
- ^ Meinhardt H., Gierer A. (2000). "Hình thành mô hình bằng cách tự kích hoạt cục bộ và ức chế bên" (PDF) . BioEssays . 22 : 753 Ảo760. doi: 10.1002 / 1521-1878 (200008) 22: 8 <753::aid-bies9> 3.0.co; 2-z. Đã lưu trữ (PDF) từ bản gốc vào ngày 2017-10-27.
- ^ Sprinzak D.; et al. (2010). "Tương tác giữa Notch và Delta tạo ra các trạng thái báo hiệu loại trừ lẫn nhau". Thiên nhiên . 465 : 86 Ảo90. doi: 10.1038 / thiên nhiên08959. PMC 2886601 . PMID 20418862.
- ^ Carlson, B.M. (2007) Nguyên tắc của sinh học tái sinh. Nhà xuất bản Học thuật, Burlington MA.
- ^ Bosch T.C.G. (2007). "Tại sao polyp tái sinh và chúng tôi không: Hướng tới một khung tế bào và phân tử để tái sinh Hydra". Sinh học phát triển . 303 : 421 Hay433. doi: 10.1016 / j.ydbio.2006.12.012. PMID 17234176.
- ^ Reddien P.W., Alvarado A.S. (2004). "Nguyên tắc cơ bản của tái sinh hành tinh". Đánh giá hàng năm về sinh học tế bào và phát triển . 20 : 725 doi: 10.1146 / annurev.cellbio.20.010403.095114. PMID 15473858.
- ^ Wagner D.E.; Vương I.E.; Reddien P.W. (2011). "Neoblasts Clonogen là tế bào gốc trưởng thành đa năng làm nền tảng cho sự tái sinh của hành tinh". Khoa học . 332 : 811 Chân816. doi: 10.1126 / khoa học.1203983. PMC 3338249 . PMID 21566185.
- ^ Nakamura T.; et al. (2008). "Phân tích tái sinh chân côn trùng thông qua can thiệp RNA". Khoa học đời sống tế bào và phân tử . 65 : 64 Tốt72. doi: 10.1007 / s00018-007-7432-0.
- ^ Simon A., Tanaka E.M. (2013). "Tái sinh limb". Nhận xét liên ngành của Wiley: Sinh học phát triển . 2 : 291 Từ 300. doi: 10.1002 / wdev.73.
- ^ Slack, J.M.W. (2013) Sinh học phát triển thiết yếu. Chương 20. Wiley-Blackwell, Oxford.
- ^ Jungnickel M.K.; Sutton K.A.; Florman H.M. (2003). "Ban đầu: bài học từ thụ tinh ở chuột và giun". Tế bào . 114 : 401 Điện404. doi: 10.1016 / s0092-8674 (03) 00648-2.
- ^ Steven, D.H. (ed.) (1975) So sánh vị trí. Nhà xuất bản Học thuật, Luân Đôn
- ^ Kimelman D., Martin B.L. (2012). "Tạo mẫu trước-sau trong phát triển ban đầu: ba chiến lược". Nhận xét liên ngành của Wiley: Sinh học phát triển . 1 : 253 Ảo266. doi: 10.1002 / wdev.25. PMC 5560123 . PMID 23801439.
- ^ Slack J.M.W. (1987). "Độ dốc hình thái – quá khứ và hiện tại". Xu hướng trong khoa học sinh hóa . 12 : 200 Linh204. doi: 10.1016 / 0968-0004 (87) 90094-6.
- ^ Rogers K. W., Schier A. F. (2011). "Gradogen Morphogen: Từ thế hệ đến giải thích". Đánh giá hàng năm về sinh học tế bào và phát triển . 27 : 377 Ảo407. doi: 10.1146 / annurev-cellbio-092910-154148.
- ^ Dahmann C.; Oates A. C.; Thương hiệu M. (2011). "Hình thành ranh giới và duy trì trong phát triển mô". Nat Rev Genet . 12 : 43 Hàng55. doi: 10.1038 / nrg2902.
- ^ Hardin J., Walston T. (2004). "Các mô hình hình thành: cơ chế và cơ chế sắp xếp lại tế bào". Ý kiến hiện tại về Di truyền & Phát triển . 14 : 399 Linh406. doi: 10.1016 / j.gde.2004.06.008.
- ^ Hammerschmidt M., Wedlich D. (2008). "Độ bám dính được điều chỉnh như một động lực của các phong trào tập trung". Phát triển . 135 : 3625 Từ3641. doi: 10.1242 / dev.015701.
- ^ O'Farrell, P. H. (2003). Làm thế nào metazoans đạt đến kích thước đầy đủ của họ: lịch sử tự nhiên của bigness. Trong tăng trưởng tế bào: Kiểm soát kích thước tế bào, (ed. MN Hall, Raff, M., và Thomas, G. (eds)), trang 1-21: Nhà xuất bản phòng thí nghiệm Cold Spring Harbor
- ^ Moss EG , Romer-Seibert J. (2014). "Thời gian nội tại tế bào trong phát triển động vật". Nhận xét liên ngành của Wiley: Sinh học phát triển . 3 : 365 trừ377. doi: 10.1002 / wdev.145.
- ^ Tata J.R. (1996). "Biến thái lưỡng cư: một mô hình tinh tế để điều chỉnh nội tiết tố của sự phát triển sau sinh ở động vật có xương sống". Dev. Tăng trưởng Diffn . 38 : 223 Điện231. doi: 10.1046 / j.1440-169x.1996.t01-2-00001.x.
- ^ Brown D.D., Cai L. (2007). "Biến thái lưỡng cư". Sinh học phát triển . 306 : 20 Hàng33. doi: 10.1016 / j.ydbio.2007.03.021.
- ^ Cohen, S.M. (1993) Phát triển đĩa tưởng tượng. Trong Bate và Martinez-Arias (chủ biên.), Sự phát triển của Drosophila melanogaster, Cold Spring Harbor Press
- ^ Maves L., Schubiger G. (2003). "Transdetermination trong đĩa tưởng tượng Drosophila: một mô hình để hiểu về tính đa năng và duy trì gen chọn lọc". Ý kiến hiện tại về Di truyền & Phát triển . 13 : 472 Bóng47. doi: 10.1016 / j.gde.2003.08.006.
- ^ Bäurle, I; Laux, T (2003). "Mô phân sinh đỉnh: Đài phun nước của tuổi trẻ". BioEssays . 25 (10): 961 Điêu70. doi: 10.1002 / b.10.10341. PMID 14505363. Đánh giá.
- ^ Leopold, AC Tăng trưởng và phát triển thực vật trang 183. (New York: McGraw-Hill, 1964).
- ^ Thương hiệu, U; Hobe, M; Simon, R (2001). "Các lĩnh vực chức năng trong mô phân sinh thực vật". BioEssays . 23 (2): 134 Công41. doi: 10.1002 / 1521-1878 (200102) 23: 2 <134::AID-BIES1020> 3.0.CO; 2-3. PMID 11169586. Đánh giá.
- ^ Barlow, P (2005). "Xác định tế bào theo khuôn mẫu trong mô thực vật: Phloem thứ cấp của cây". BioEssays . 27 (5): 533 Điêu41. doi: 10.1002 / bies.20214. PMID 15832381.
- ^ Ross, S.D.; Pharis, R.P.; Binder, W.D. 1983. Chất điều hòa sinh trưởng và cây lá kim: sinh lý học và tiềm năng sử dụng của chúng trong lâm nghiệp. tr. 35 Chân78 trong Nickell, L.G. (Ed.), Hóa chất điều hòa sinh trưởng thực vật. Tập 2, CRC Press, Boca Raton FL.
- ^ Jin JP; et al. (Tháng 7 năm 2015). "Một bản đồ điều hòa phiên mã Arabidopsis cho thấy các đặc điểm chức năng và tiến hóa riêng biệt của các yếu tố phiên mã mới". Sinh học phân tử và tiến hóa . 32 (7): 1767 Điêu1773. doi: 10.1093 / molbev / msv058. PMC 4476157 . PMID 25750178. Được lưu trữ từ bản gốc vào ngày 2016-06 / 02.
- ^ Nieuwkoop, P.D. và Faber, J. (1967) Bảng bình thường của Xenopus laevis (Daudin). Bắc Hà Lan, Amsterdam.
- ^ Harland R.M., Grainger R.M. (2011). "Nghiên cứu Xenopus: biến chất bởi di truyền và genomics". Xu hướng di truyền học . 27 : 507 Ảo515. doi: 10.1016 / j.tig.2011.08.003. PMC 3601910 . PMID 21963197.
- ^ Lawson N. D., Wolfe S. A. (2011). "Phương pháp tiếp cận di truyền ngược và ngược để phân tích sự phát triển của động vật có xương sống ở Zebrafish". Tế bào phát triển . 21 : 48 trận64. doi: 10.1016 / j.devcel.2011.06.007.
- ^ Hassan Rashidi V.S. (2009). "Phôi gà: ấp nở một mô hình cho nghiên cứu y sinh đương đại". BioEssays . 31 : 459 cường65. doi: 10.1002 / bies.200800168.
- ^ Behringer, R., Gertsenstein, M, Vintersten, K. và Nagy, M. (2014) Thao tác với phôi chuột. Một hướng dẫn thí nghiệm, phiên bản thứ tư. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press.
- ^ St Johnston D (2002). "Nghệ thuật và thiết kế màn hình di truyền: Drosophila melanogaster". Nat Rev Genet . 3 : 176 Từ188. doi: 10.1038 / nrg751. PMID 11972155.
- ^ Riddle, D.L., Blumenthal, T., Meyer, B.J. và Priess, J.R. (1997) C.elegans II. Nhà xuất bản Phòng thí nghiệm Cold Spring Harbor, Cold Spring Harbor, NY.
- ^ Ettensohn C.A., Sweet H.C. (2000). "Tạo phôi phôi nhím biển sớm". Curr. Hàng đầu. Nhà phát triển Biol . 50 : 1 Đấu44.
- ^ Lemaire P (2011). "Ngã tư tiến hóa trong sinh học phát triển: áo dài". Phát triển . 138 : 2143 Lỗi2152. doi: 10.1242 / dev.036175. PMID 21558365.
- ^ Nacu E., Tanaka E.M. (2011). "Tái sinh Limb: Một sự phát triển mới?". Đánh giá hàng năm về sinh học tế bào và phát triển . 27 : 409 trừ440. doi: 10.1146 / annurev-cellbio-092910-154115.
- ^ Reddien P.W., Alvarado A.S. (2004). "Nguyên tắc cơ bản của tái sinh hành tinh". Đánh giá hàng năm về sinh học tế bào và phát triển . 20 : 725 doi: 10.1146 / annurev.cellbio.20.010403.095114. PMID 15473858.
- ^ Ader M., Tanaka E. M. (2014). "Mô hình hóa sự phát triển của con người trong văn hóa 3D". Ý kiến hiện tại về sinh học tế bào . 31 : 23 Hàng28. doi: 10.1016 / j.ceb.2014.06.013.
- ^ Weigel, D. và Glazebrook, J. (2002) Arabidopsis. Hướng dẫn thí nghiệm. Nhà xuất bản Phòng thí nghiệm Cold Spring Harbor, Cold Spring Harbor, NY.
Đọc thêm [ chỉnh sửa ]
- Gilbert, S. F. (2013). Sinh học phát triển . Sunderland, Mass.: Sinauer Associates Inc.
- Slack, J. M. W. (2013). Sinh học phát triển thiết yếu . Oxford: Wiley-Blackwell.
- Wolpert, L. và Tickle, C. (2011). Nguyên tắc phát triển . Oxford và New York: Nhà xuất bản Đại học Oxford.
Liên kết ngoài [ chỉnh sửa ]